Кондилартите - древните копитни бозайници.
 |
Филогения на Condylarthra. |
Разредът Condylarth
ra е една от най-характерните групи бозайници за палеоцена. И илюстрира добре еволюционното ниво на фауната през този период. В сравнение с днешните бозайници кондилартите са сравнително неспециализирани плацентни бозайници. Но в сравнение със своите насекомоядни предшественици, те показват първи признаци на всеядство и дори на растителноядство.
Заради отсъствието на сушата на едри растителноядни след изчезването на динозаврите, тази особеност в диетата дава началото на бързата еволюция и разпространение на кондилартите, което се наблюдава през Палеоцен. В резултат на това се появяват различни групи унгулати (или копитни бозайници), които формират доминиращата растителноядна фауна в повечето неозойски животински съобщества на сушата, с изключение на островния континент Австралия. Терминът „унгулати” произхожда от наименованието на подгрупа плацентни бозайници (Ungulata), които са потомци на същия предшественик – най-примитивния кондиларт. От съвременните бозайници, чифтокопитни и нечифтокопитни, дамани, слонове, тръбозъби, морски крави и китове се причисляват към надразред Ungulata (виж края на статията). Освен кондилартите още няколко изчезнали групи би следвало да се причислят към унгулатите, особено ендемичните южноамерикански разреди. Въпреки че много от членовете на групата имат копита, тази особеност не характеризира групата като цяло. Някои кондиларти наистина имат малки копитца, но повечето примитивни видове са с нокти. От друга страна, копита са притежавали животни, които не могат да бъдат причислени към Ungulata, като изчезналите
Pantodonta.
Повечето кондиларти са познати от Северна Америка, континентът с най-богати фосилни находки на палеоценски бозайници. Само няколко палеоценски фауни са открити в Европа, но находките показват голямото значение на кондилартите и в тази част на света, а представителите им са били близкородствени на северноамериканските кондиларти. Изненадващо само два все още спорни
вида кондиларти са описани от иначе богатата палеоценска фауна на Азия. Екологичната роля на растителноядни бозайници там е заета от Anagalida и Pantodonta. Същите са ставали жертва на месоядните потомци на кондилартите, разред
Mesonychia. Южна Америка е единственият континент в южното полукълбо, който има съществени находки на палеоценски бозайници, а нарастващият брой южноамерикански кондиларти ги прави важен компонент на фауната.
Най-примитивният известен кондиларт е плъхоподобният
Protoungulatum, намерен в С.Америка и Канада. Освен разположението на останките му в много ранни пластове, животни от този род са намирани и заедно със зъби на динозаври. По тази причина първоначално се е смятало, че
Protoungulatum са се срещали през късна Креда, съществували са заедно с последните динозаври и типичните кредни бозайници в т.нар. съобщества от Бъгкрийк. При това положение ранните унгулати като
Protoungulatum трябва да са били в конкуренция с растителноядните динозаври и вероятно са допринесли за изчезването им. По-съвременната интерпретация представя находките от Бъгкрийк като смесени раннопалеоценски и къснокредни фосили. Къснокредните вероятно са ерозирали и отнесени от реката, а впоследствие отложени върху пресни раннопалеоценски пластове. Решението не е окончателно, но новата интерпретация отговаря на идеята, че бозайниците са запълнили освободените екологични ниши след катастрофичното изчезване на динозаврите, което може би е причинено от сблъсък на голям метеорит. Но така или иначе прецизното датиране на фосилите и статуса на
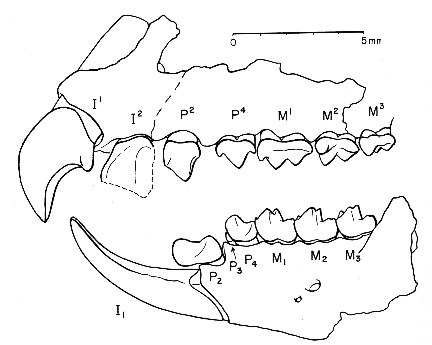
Protoungulatum като прототип на по-късните унгулати не е оспорван откакто са първоначално описани през 1965г. Макар и примитивни, в зъбната редица на
Protoungulatum се наблюдават сходства с типичните черти, наблюдавани при унгулатите: ниски зъби със способност за надробяване и смилане, което вероятно е позволявало на животните да се хранят с по-фина растителност, плодове, но и някои насекоми. Само няколко кости са намерени от тези най-ранни кондиларти, освен техни челюсти и зъби. Голям прогрес е, че потенциален предшественик на
Protoungulatum е бил идентифициран в къснокредни пластове в Узбекистан. Тези малки плацентни бозайници, наречени целестиди, не са окончателно класифицирани като унгулати, но зъбите им показват първите етапи от еволюцията на унгулатоподобна зъбна редица.
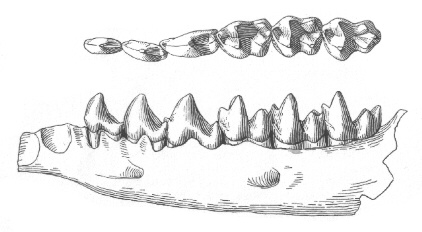
Protoungulatum традиционно се смятат за членове на сем. Arctocyonidae, което се развива и достига разцвет през Палеоцен в С.Америка и Европа. Арктоционидите са най-малката група растителноядни кондиларти. Всъщност техните скелети са изглеждали до голяма степен като тези на месоядните, с големи и сравнително остри кучешки зъби, но арктоционидите не са притежавали специализирани зъби за разрязване на месото и е по-вероятно да са били всеядни. Крайниците им са били относително къси и не показват нито една от специализациите, които обикновено свързваме с унгулатите, като редукция на крайните пръсти, срастване на костите или наличие на копита. Повечето арктоциониди са познати единствено от техните зъби, които показват много индивидуални вариации, така че систематиката на различните родове арктоциониди е незавършена в момента.
 |
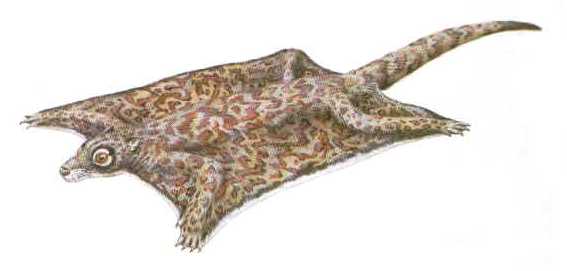
| Реконструкция на Chriacus. |
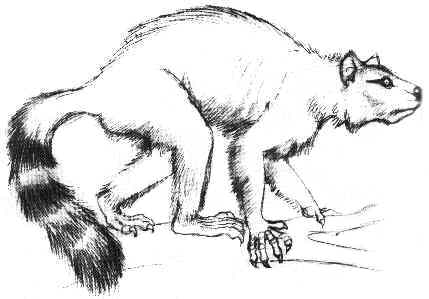
Със телесно тегло от 5 до 10 кг.
Chriacus е типичен пример за малък арктоционид. Почти пълен скелет е открит в ранноеоценски пластове в Уайоминг. Той е наистина примитивен по структура и подобен на раннопалеоценските арктоциониди като
Loxolophus, които са малко по-добре изучени. Както показва този скелет,
Chriacus е бил еднакво пригоден за живот по дърветата и на земята. От съвременните животни може да бъде най-добре оприличен на ракуните и циветите. Както повечето катерещи се животни
Chriacus притежава мощна мускулатура на крайниците, много подвижни стави и пригодени за хващане на храна петопръсти предни крайници. Опашката на
Chriacus е дълга и груба, използвана е за балансиране, възможно е да е била снабдена и с хватателна част. Както показва формата на предните крайници, животните са били приспособени за копаене.
Chriacus вероятно са се хранили с плодове, насекоми и други дребни животни. Интересно е, че горните им резци са образували нещо като гребло, което вероятно е използвано за дълбаене.
Можем да предположим, че едрите арктоциониди, като
Arctocyon и
Claenodon, въпреки че също са били способни да се катерят по дърветата, са прекарвали по-голямата част от живота си на земята. Екологично тези животни могат да се сравнят с мечките. Възможно е периодично да са заемали ролята на едри хищници в палеоценската фауна, заедно с други две семейства унгулати, еволюирали от ранните арктоциониди – трисодонтидите и мезонихидите. Последните две групи са класифицирани също и като кондиларти, но сега се считат за членове на разред Mesonychia. Всички тези месоядни унгулати се разглеждат като отделно звено на ранните хищни бозайници.
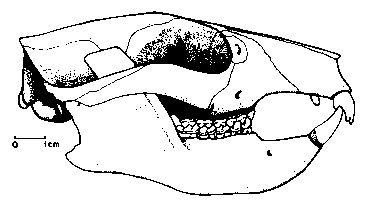
Освен арктоционидите, членовете на сем. Periptychidae са били доминиращите кондиларти в раннопалеоценската фауна на С.Америка. По-малко от 1 млн. години след края на Креда те вече са развили разнообразни размери, от катерицоподбните
Anisonchus, до
Ectoconus с размерите на овца, един от най-едрите бозайници на Пуеркан. Пълен скелет на
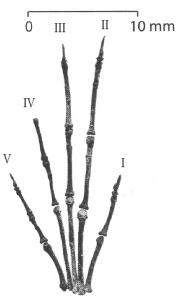
Ectoconus ditrigonus бе открит в Ню Мексико, правещ този вид най-добре изучения бозайник от фауната на Пуеркан. Една от лакътните стави на скелета е увредена, което вероятно означава, че животното е било куцо. Общата телесна структура на
Ectoconus е подобна в много отношения на тази на едрите арктоциониди – като масивна конструкция, малка мозъчна кутия на черепа, силни и къси крайници и дълга силна опашка. Петте широкоразположени пръсти на крайниците им са имали малки копитца, подобни на тези на днешните тапири, за разлика от
завършващите с нокти пръсти на арктоционидите. Въпреки че
Ectoconus почти идеално се доближават по форма на тялото до унгулатите, зъбната редица на периптихидите е специализирана по особен начин, който ги изключва като възможни предшественици на съвременните унгулати. Повечето периптихиди имат уголемени, широки предкътници, които вероятно са служили за справяне с груба растителна храна. Силни бразди се спускат от върховете на тези зъби, намерени в последният оцелял род къснопалеоценския
Periptychus, животно малко по-дребно от
Ectoconus, но с по-голяма глава. Характерните предкътници на
Periptychus са много подобни на тези на някои прасета и всъщност тези животни са имали диета много сходна с тази на прасетата или пекаритата.
 |
| Реконструкция на Ectoconus. |
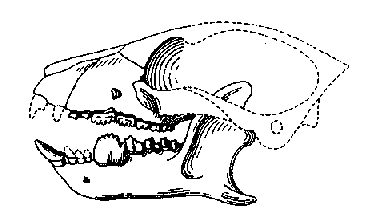
Трето семейство от кондиларти – Hyopsodontidae – добива значение в късен Палеоцен, когато периптихидите вече са минали зенита на своята еволюция. Хиопсодонтидите са били типични дребни животинки, имащи много малко общо с представата ни за унгулатите и са приличали повече на насекомоядните по външен вид. В някои случаи сходството с насекомоядните включва и такива детайли като зъбите и съществуват предположения, че някои хиопсодонтиди, като
Litomylus, са всъщност насекомоядни, родствени на таралежите. Най-добре проученият хиопсодонтид е
Hyopsodus, много успешен род, срещал се е в ранен Еоцен в цялото северно полукълбо и е един от последните кондиларти – разбира се, това е и въпрос на дефиниране на групата. Няколко фосилни находки от С.Америка показват, че
Hyopsodus за първи път се появява там през късен Палеоцен.
Hyopsodus е дребно животно, с размерите на плъх, с къси крайници, завършващи с нокти. Скелетът му показва, че животното е живеело частично и върху дърветата. Европейското подсемейство на хиопсодонтидите е от къснопалеоценския период на Германия и Франция. Родът
Pacshatherium вероятно е бил многоброен през късен Палеоцен и ранен Еоцен в Европа, тъй като на места съставлява значителна част от бозайниковите фосили. Глезенните кости на тези животни показват, че
Pacshatherium вероятно са били пъргави по дърветата, сравними с днешните катерици.
 |
| Реконструкция на Hyopsodus. |
Mioclaenus и други техни родственици от Палеоцена на С.Америка дълго време са считани като друго подсемейство на хиопсодонтидите. Днес те се разглеждат като отделно семейство на кондилартите - Mioclaenidae. Миокленидите са били малко по-едри от хиопсодонтидите – с размерите на заек. Уголемените и широки предкътници са типични и за миокленидите, също както при периптихидите. Те вероятно са подпомагали преработката на твърда, фиброзна растителност. Среднопалеоценският род
Mioclaenus притежава най-уголемените предкътници.
Не са известни миоклениди от Европа, а
Pleuraspidotherium от късния Палеоцен на Франция вероятно погрешно е включен в това семейство.
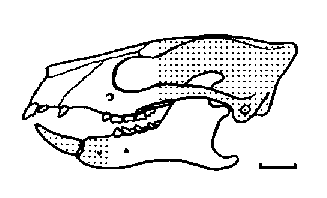
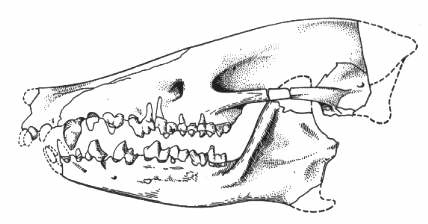
Pleuraspidotherium, животно с дължина на тялото около 60см., е един от най-често срещаните бозайници в речните наноси около Церн.
Pleuraspidotherium е притежавал изумително развити дъвкателни зъби за палеоценски бозайник, сходни с тези при някои съвременни чифтокопитни, като елените и камилите. Снабдени с подобна зъбна редица,
Pleuraspidotherium вероятно са били най-успешните растителноядни животни за времето си. Скелетът им е сходен с този на арктоционидите, но без адаптациите за катерене по дърветата на последните.
 |
| Череп на Pleuraspidotherium aumonieri. |
Дълго време фосили на миоклениди са били намирани само в северното полукълбо, но нови открития напоследък показват, че тези кондиларти са колонизирали и Ю.Америка, при това доста успешно. До сега в Ю.Америка са открити само миоклениди от ранен Палеоцен, като
Molinodus и
Tiuclaenus, локализирани в Тиупампа, Боливия.
Едно ендемично за Ю.Америка семейство кондиларти, Didolodontidae, е от по-късен период. Дидолодонтидите и миокленидите имат много сходни зъбни редици, което подсказва, че дидолодонтидите са еволюирали от миокленидите в Ю.Америка. За съжаление фосилните находки от дидолодонтиди са недостатъчни, като зъби, челюсти и отделни глезенни кости. Били са предимно дребни животинки, като
Ernestokokenia от късен Палеоцен до ранен Еоцен в Аржентина, но са включвали и по-едри видове като среднопалеоценските
Lamegoia от Бразилия. По отношение на зъбната си редица дидолодонтидите са почти неразличими от най-примитивните представители на Litopterna, важен разред ендемични южноамерикански унгулати, от които по-късно произлизат конеподобните и камилоподобни форми. Важна разлика с дидолодонтидите е наличието при литоптерните на по-развити крайници, особено в областта на глезена. Тези промени вероятно са свързани с подскачащият начин на придвижване на ранните литоптерни, което позволява на по-късните видове да се приспособят към бързо бягане. Свидетелствата от зъбната редица показват недвусмислено, че също както и дидолодонтидите, литоптерните произлизат от миокленидите. За да се представи по-точно тази роднинска връзка, наскоро бе предложено да се отдели нов разред бозайници – Panameriungulata – за миокленидите, дидолодонтидите и литоптерните. Това би било подходящо място и за систематизиране на примитивни форми с неустановена глезенна структура, които не могат със сигурност да се определят като дидолодонтиди или литоптерни, като
Asmithwoodwardia, много дребно животинче от среден Палеоцен на Бразилия и ранен Еоцен на Аржентина.
Обратно в северното полукълбо, друго семейство кондиларти, Phenacodontidae, вероятно включва предшествениците на един много по-познат разред унгулати – нечифтокопитните Perissodactyla, представени от конете, носорозите и тапирите в днешната фауна. Исторически фенакодонтидите формират „ядрото” на Condylarthra. Намерени са добре запазени скелети на рода
Phenacodus, който е добър пример за прародител-унгулат с начални адаптации за бързо бягане. За разлика от арктоционидите, периптихидите и миокленидите, фенакодонтидите не са част от първата вълна кондиларти, населили С.Америка. Те са представени на този континент първо с подобните на лисица по размери
Tetraclaenodon в средата на Палеоцен. Наличието на по-напреднали фенакодонтиди като
Phenacodus и
Ectocion бележат началото на късен Палеоцен в С.Америка. Родът
Phenacodus включва разнообразни по размери фенакодонтиди, включително и животни с размери подобни на днешните овце. Членовете на род
Ectocion обикновено са по-малки, с телесно тегло от само три килограма при най-дребните видове, като се наблюдава известно застъпване в това отношение между двата рода. Фенакодонтидите са били доминиращите бозайници в С.Америка през късен Палеоцен и са съставлявали повече от 50% от видовете в тогавашната фауна на континента.
Phenacodus и
Ectocion съществуват до средата на Еоцен, като към края на Палеоцен числеността на фенакодонтидите намалява. Интересно изключение представлява локалното изобилие на фенакодонтидът с размерите на куче –
Meniscotherium, който на места е формирал истинско струпване на фосили.
Meniscotherium е датиран основно като ранноеоценски, въпреки че най-ранните представители вероятно са съществували още в края на Палеоцен.
Phenacodus се разпространяват в Европа в началото на Еоцен по времето на голям фаунистичен обмен между Новия и Стария свят, но никога не става така съществен компонент на европейската фауна.
 |
| Реконструкция на Phenacodus. |
Скелетите на фенакодонтидите са твърде примитивни, особено по отношение на дългата и тежка опашка, но някои сходства с перисодактилите се наблюдават при
Phenacodus: крайниците му са по-дълги, отколкото при по-примитивните кондиларти и завършват с пет пръста с копита, като първият и петият пръст са с редуцирана дължина. Това предшества същата особеност при ранноеоценските коне
Hyracotherium (понякога наричани „
Eohippus”), които пък напълно са загубили първия пръст на предния си крайник, и първия и последния пръст на задния. Това устройство показва, че
Phenacodus до голяма степен са били добре приспособени за бягане. Известни са само няколко останки на крайници от
Ectocion, но те показват, че тези дребни животни са били в известна степен по-добри бегачи от по-едрите си родственици. Черепът на фенакодонтидите е удължен и с малка мозъчна кутия. При
Phenacodus intermedius назалните кости са удължени като при днешните тапири, което показва, че тези животни са притежавали къс хобот. От зъбната редица може да се съди, че по-късните фенакодонтиди са били растителноядни: техните дъвкателни зъби са имали малки вдлъбнатини, които понякога са се сливали в ръбове, подобно на зъбите на ранните перисодактили като
Hyracotherium. При
Meniscotherium тези гънки образуват полулунна форма, което донякъде изглежда като преждевременно развита адаптация за раннотерциерски бозайник, която може да бъде сравнена само със зъбната редица на
Pleuraspidotherium. Ранните изследвания ги определят като родствени групи, но по-късни проучвания показват, че не съществува такава връзка между
Pleuraspidotherium и
Meniscotherium, а последният е всъщност най-високоспециализираният фенакодонтид.
По-ранни изследвания показаха, че разред Condylarthra включва форми, които са били адаптирани към разнообразни условия на живот, като катерицоподобни, катерещи се по дърветата животни, растителноядни с размерите на днешни овце и големи подобни на мечки всеядни. Въпреки че всички кондиларти имат общ предшественик, близък до
Protungulatum, те са обединени от систематиката предимно въз основа на това, че са по-примитивни в сравнение с по-развитите унгулати. Поради тази причина разредът често е критикуван като „кошче за боклук” за примитивните форми унгулати и е предложено отхвърлянето му. Няколко други разреда са били предложени да включат членовете на Condylarthra, най-вече Panameriungulata за разклонението, което води до южноамериканските литоптерни, но досега няма постигнат консенсус по този въпрос.
От друга страна трябва да отбележим, че ролята на кондилартите като предшественици на унгулатите, не е напълно документирана и подкрепена от достатъчно фосили. Може да се каже, че в този случай търсенето на „липсващо междинно звено” продължава. Litopterna е група от унгулати, които с по-голяма сигурност могат да се прибавят към друга, по-специфична група кондиларти, миокленидите. Случаят със сходноадаптираните, нечифтокопитни унгулати, или Perissodactyla, в северното полукълбо е по-неясен. В случай, че се докаже, че те наистина са еволюирали от фенакодонтидите, то тогава трябва да бъдат отделени от по-примитивните форми като
Tetraclaenodon, но междинни форми все още не са открити. Интересен череп с впечатляващи перисодактилни зъби наскоро беше открит в Китай в къснопалеоценски пластове и именуван
Radinskya. Въпреки че не са открити останки от крайници на
Radinskya, това животно представлява правдоподобен вариант за произхода на нечифтокопитните унгулати. Родствените връзки на
Radinskya с останалите палеоценски бозайници не са съвсем ясни, но е възможно да принадлежи към ендемичното азиатско семейство растителноядни, наречени Phenacolophidae.
Молекулярните изследвания поставят под въпрос традиционните схващания. Молекулярните биолози вече преместиха слоновете, даманите, тръбозъбите и морските крави от Ungulata в нова група – Afrotheria, африканско разклонение плацентни бозайници, което включва още тенреците, златните къртици и слонските земеровки. Ако това бъде потвърдено и от бъдещи изследвания, съвременните унгулати, тези съществуващи и днес наследници на Condylarthra, ще бъдат ограничени до нечифтокопитните, чифтокопитните и китовете.
Съвременните изследвания на части от скелета на друг кондиларт показват, как едно изненадващо откритие може да възроди идеи, които са били отхвърлени. Съдейки по зъбите, екземплярът от срадния Палеоцен на Ню Мексико представлява малък арктоционид, подобен на
Chriacus. Освен адаптациите за катерене по дърветата, характерни и за
Chriacus, фрагментите от крайници на новия екземпляр показват начални адаптации и за бягане, удивително сходни с тези при ранните представители на чифтокопитните унгулати Artiodactyla. Това представлява ново доказателство в подкрепа на хипотезата, първоначално базирана на факти от зъбните редици, че Artiodactyla са потомци на арктоционидите. В същото време това изненадващо откритие показва, че палеоценските бозайници са били много по-разнообразни в адаптациите си, отколкото техните зъби и челюсти могат да ни покажат.