До 1980 г. фосилните находки от ранни четиритоги са били ограничени в рамките на Ichthyostega - къснодевонски род, известен от находища в източната част на Гренландия. Но след 1987 г. находките се увеличават драстично, както и такива на девонски (фаменски) риби, предшественици на четириногите. Тези находки заедно с откритията в други научни дисциплини са причина за едно ново, по-съвременно разбиране за еволюцията на четириногите. Вместо да се разглежда преминаването от
водни риби към
сухоземни четириноги, с новия подход се разглеждат и
двата процеса - прехода от риби към четириноги и преминаването от водна към сухоземмна среда.
Едни от най-важните находки, имащи отношение към новото разбиране за еволюцията на четириногите, са тези на Acanthostega от края на 80-те и началото на 90-те години на миналия век. Покривни кости от череп на Acanthostega са открити още през 1933 г. от Гунар Содърбърг и Ерик Яврик, но важността им не е отчетена преди откриването на допълнителните находки през 1987г. от Джени Кларк. Заедно с Майкъл Коутс те установяват, че това е едно очевидно четириного животно, което обаче не е достатъчно, за да бъде определено като сухоземно. Крайниците му са били подходящи единствено за да поддържат част от собственото тегло, а китки отсъстват. Въпреки това се наблюдават добре развити пръсти - на предни и задни крайници. Изненадващо предните крайници притежават осем пръста (вместо очакваните пет), а задните седем. Допълнителни характеристики на гръбначния стълб, ребрата, таза и опашката потвърждават идеята, че Acanthostega не биха били в състояние да поддържат теглото си на сушата. Възможно е подобно на днешните делфини и китове Acanthostega вторично да са изгубили скелетните характеристики необходими за живот на сушата. Тази възможност обаче е в противоречие с наличието на вътрешни рибоподобни хриле (дишат също така и с бял дроб). Acanthostega със своите крайници, характеристики на таза и различни други функции е едно типично живонто, което не е било в състояние да напуска водната среда.
Неочакваните характеристики на Acanthostega са довели до преразглеждането на род Ichthyostega от Пер Алберг, Дженифър Кларк и Майкъл Коутс. Предните крайници на Ichthyostega са по-груби от тези на Acanthostega и изглежда са били в състояние да поддържат предната половина на тялото. Задните крайници обаче вероятно са функционирали повече като гребла, отколкото като крака. Подобно на Acanthostega са притежавали седем пръста на задните си крайници, но няма следи от вътрешни хриле.
Работата по Acanthostega и последвалите изследвания на Ichthyostega подновяват интереса и към други къснодевонски четириноги. Това става повод за преразглеждане на вече известни групи, например Tulerpeton, както и за откриването на нови такива, например Hynerpeton. Освен това по-доброто разбиране на анатомията на ранните четириноги улеснява повторната оценка на разнообразни музейни екземпляри, първоначално определени като ръкоперки, например Elginerpeton и Ventastega. Находките от тези четириноги са значително по-непълни от тези на Acanthostega и Ichthyostega. От тях Tulerpeton и Ventastega са най-добре представени. Останалите обикновено са представени от един или само няколко отделни елемента, въпреки че някои от тези елементи могат да бъдат особено информативни.
Следствие от новите находки от къснодевонски четириноги е и по-доброто разбиране на палеонтолозите за еволюцията на техните най-близкородствени риби - Panderichthys и Elpistostege. Тези къснодевонски риби споделят редица характеристики с ранните четириноги. В действителност изкопаем череп на един от тези видове, Elpistostege, първоначално е бил класифициран като четириного, макар че се различава от тях по ключови характеристики. Съвсем наскоро (2006г.) откритието на друга четириногоподобна риба, Tiktaalik, значително обогатява разбиранията за прехода от ръкоперки към четириноги. Този вид показва характеристики, които са
междинни за Panderichthys и къснодевонските четириноги. Времевият период между тези четириногоподобни риби и четириноги като Acanthostega и Ichthyostega е около 10-15 млн. години. Но има една празнина във фосилните записи от около 15 млн. години след края на Фамен, след която се наблюдава изключително разнообразие от находки на водни и сухоземни четириноги.
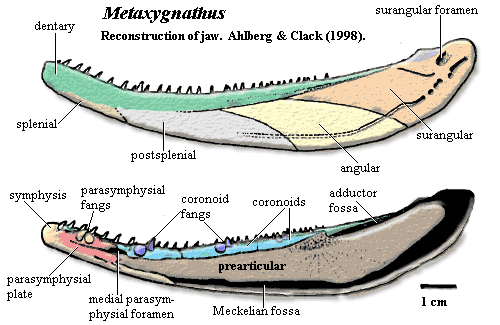
Най-ранните останки на четириноги са открити в Еврамерика - палеоконтинент, състоящ се от днешните Северна Америка, Прибалтика, Северна Англия и Европейска Русия. Изключение са находките от Австралия - Metaxygnathus - която е била локализирана в другия край на Гондвана, както и тези от Китай - Sinostega - разположен по това време на север от източната част на Гондвана. Дали това биогеографско разпространение е резултат на пристрастност при взимане на пробите, или има други причини, засега не е известно. Но всички местообитания на ранни четириноги се характеризират с тропичен до субтропичен климат. Освен това, с изключение на Tulerpeton и вероятно Jakubsonia, които са открити в естуарни депозити, всички къснодевонски четириноги, както и фаменските риби Panderichthys и Elpistostege, са обитавали сладководни басейни. Тези сладководни басейни и околните им пейзажи са претърпели значителна трансформация. Суровата, неплодородна и нестабилна сладководна среда, както и сухоземните местообитания през среден Силур дават път на онези видове, които се адаптират към появата и еволюцията на висшите растения.

Черепни покривни кости от Acanthostega gunneri са открити за първи път във фаменски депозити от източната част на Гренландия (на 360 млн. години) през 1933 г., описани и именувани през 1952 г. от Ерик Яврик. Допълнителни фосилни останки са открити по време на геоложка експедиция през 1970 г., но те потъват в неизвестност, преди да бъдат преоткрити от Дженифър Кларк. По време на експедиция водена от Кларк и Алберг през 1987 г. са намерени няколко изключително добре запазени екземпляра. Изобилието и качеството на останките от Acanthostega ги правят едни от най-добре проучените ранни четириноги. След изследването на тези останки Кларк и Коутс съобщават за няколко забележителни находки, които налагат промяна в разбирането ни за ранните четириноги. Преди тези находки повечето учени приемат, че развитието на крайниците и ходилата е провокирано от излизането на сушата. Кларк и Коутс обаче описват едно от най-ранните четириноги, което е било неспособно за живот на сушата. То има добре обособени пръсти (на предни и задни крайници), но не и китки или глезени. Има сравнително дълги кости на крайниците, които обаче не са способни да поддържат голяма тежест. Тазът също не е бил способен да поддържа голямо тегло, тъй като е бил не така здраво прикрепен към гръбначния стълб. Здравото прикрепване всъщност не би помогнало така или иначе, доколкото структурата на гръбначния стълб се е базирала основно на нотохордата, а не на поредица от взаимносвързани, гъвкави прешлени. Гръбнакът е бил добре приспособен за справяне с механичното натоварване при плуване, но почти безполезен за поддържане на тегло на сушата. Освен това тънките и къси ребра не са били в състояние да предпазват жизненоважните органи. Acanthostega са притежавали и опашка, която е била обградена с дълга костна перка. Накратко, имали са опашка подходяща за плуване, гръбначен стълб на риба и лопаткоподобни крайници.
Предимно (ако не и изключително) водният начин на живот на Acanthostega се потвърждава и от наличието на вътрешни хриле. (Свидетелства за наличие на вътрешни хриле са например костните хрилни дъги. Всички хрилнодишащи земноводни от Карбон до наши дни притежават външни хриле.) Други характеристики, потвърждаващи водния начин на живот са устройството на стремето (кост, която еволюира в средното ухо при сухоземните четириноги) и запазването на сензорната странична линия, срещаща се и при днешните риби. Малките рибоподобни ноздри на Acanthostega вероятно са били използвани за обоняние под водата, въздухът изглежда е бил транспортиран до белия дроб на глътки.
Въпреки че Acanthostega притежават много рибоподобни характеристики, те все пак са притежавали крака, а не перки. Техните крака също преобръщат разбирането ни относно еволюцията на крайниците на ранните четириноги. Ранните схващания, че първите четириноги са притежавали по пет пръста, са базирани основно на необосновани допускания и асоциации, но Acanthostega имат по осем пръста на предните, и по седем или осем пръста и на задните си крайници. Последвалите анализи показват, че най-малко още две други групи ранни четириноги, Ichthyostega и Tulerpeton, също са притежавали по повече от пет пръста.
Със своята комбинация от характеристики на риби и четириноги Acanthostega пораждат разнообразни спекулации относно палеоекологията и еволюцията на ранните четириноги. Крайниците им биха били много полезни като перки в плитки, изпълнени с водна растителност и дървесни отпадъци води. Такива плитки води могат да бъдат разнообразни влажни зони, разливи на реки или заливни терени. Acanthostega също така притежават защитна покривка от продълговати яйцевидни люспи на коремната страна, но не и на останалата част на тялото. Това вероятно е защита от абразия по време на движение.
Останките на Acanthostega са откривани в речни депозити, където вероятно са живеели. Достигали са до около 60 см. на дължина.
Densignathus rowei
Densignathus rowei е второто четириного открито в Ред Хил, Пенсилвания. Първото е Hynerpeton bassetti, както и един изолиран, несистематизиран все още екземпляр (ANSP 21350). Находката от денсигнатус представлява единична долна челюст. Тя е доста груба и притежава смесица от характеристики, някои от които предполагат по-напреднало развитие (частично затворен долночелюстен канал), докато други - по-примитивен рибоподобен статус (наличие на короноидни зъби).
Elginerpeton pacheni
Находките на Elginerpeton pancheni са намерени в Шотландия. Първоначално са били идентифицирани като "неизвестен саркоптериг", а преразглеждането им в началото на 1990 г. от Пер Алберг показва, че принадлежат на ранно четириного. Всъщност Elginerpeton и близкородствените Obruchevichthys са най-ранните четириноги, известни от фосилни находки. Останките включват фрагменти от рамо и таз, фемур, тибия и части от горна и долна челюст. Предполага се, че животните са достигали 1,5 метра на дължина.
Челюстите притежават смесица от характеристики на четириноги и къснодевонски риби. Тазът и крайниците имат сходни черти с Ichthyostega, докато раменният пояс е сходен повече с този на Hynerpeton или Tulerpeton. За съжаление не са открити крака (или рибоподобни перки). Следователно не е съвсем ясно дали Elginerpeton са по-близкородствени с Panderichthys или с другите ранни четириноги. Освен това Elginerpeton и техните съвременници Obruchevichthys притежават някои уникални характеристики, които ги отделят от фаменските четириноги. Предната част на черепа е много по-тясна, а общата му дължина е много по-голяма. Obruchevichthys и Elginerpeton вероятно представляват отделни, кратко съществували разклонения, които се отделят преди появята на останалите четириноги.