Почти всички групи големи гръбначни животни - на сушата, в моретата и въздуха – всички динозаври, плезиозаври, мозазаври и птерозаври внезапно изчезват преди около 65 млн. години, в края на креда. По същото време изчезват голяма част от планктона и много тропически безгръбначни, особено рифовите обитатели, много сухоземни растения също биват засегнати. Това грандиозно изчезване на видове бележи известната К/Т (креда-терциер) граница в историята на Земята и края на мезозойската ера. К/Т събитията са били глобални, засегнали са всички големи континенти и световния океан.
Индикатори за катастрофалните събития са внезапното изчезване на планктона и други морски организми, както и рязката промяна във видовия състав на полена в съответните пластове. Тази промяна свидетелства за внезапно изчезване на покритосеменните растения в края на креда и заместването им от папратови, а след това обратно завръщане на нормалната флора. Този папратов пик е открит в много сухоземни К/Т пластове в западните части на Северна Америка, където фосилните записи от сухоземна растителност са доста подробни и е интерпретиран като следствие на катастрофално количество прах в атмосферата: папратите се възстановяват първи, колонизират освободените площи, последвани от покритосеменните едва след възстановяването на почвите. Това заключение е направено на базата на аналогията с наблюдавани подобни обрати в областта на флората, предизвикани от вулканични изригвания. Разбира се, трябва да бъде отчетен факта, че подобни аномалии са ограничени до фосилните записи на Северна Америка. Навсякъде другаде по света, особено в южното полукълбо, не се наблюдава нищо повече от постепенна промяна във видовия състав на растенията през К/Т периода.
За оценка на степента на въздействие и евентуалните промени в екосистемите е необходимо прецизното им и точно реконструиране. Това е предмет на палеоекологията. Вероятно повечето от вас ще бъдат разочаровани да научат, че предмет на палеоекологията не са евентуални палеопротести срещу замърсяващите атмосферата отровни летливи газове на Цехщайново море, нито пък отчаяни палеодействия по съхраняването на българските мезозойски корали, страдащи от необратимите промени в климата. Палеоекологията изучава начина на живот на отделни изкопаеми организми, животни и растения, отношенията помежду им, както и със заобикалящата ги среда, и методите й играят основна роля при проучванията на масовите измирания. Това, разбира се, не винаги е лесно и възможно, като се има предвид непълнотата на фосилните записи и т. нар. ефект на Сигнор-Липс - поради непълни фосилни данни, съществува вероятност повечето измрели видове да са изчезнали реално по-късно от времето, датирано по най-младия намерен фосил.
Масовото измиране в края на креда безспорно е най-популярното от петте големи, главно заради изчезването на енигматичните динозври. До преди 1980 г. относно причините за това са предлагани най-различни хипотези. Те варират от приемливи такива (глобални климатични промени, таксономични промени в растителните съобщества, астероиден сблъсък, тектонични процеси, промени в нивото на световния океан) до по-налудничави (загуба на сексуален апетит, изглупяване и хормонален дисбаланс при динозаврите; ожесточена конкуренция с гъсеници за растителна храна; бозайници, които изяждат всички динозавърски яйца). Множество експедиции и изследвания са проведени с цел да бъде установено какво точно е причинило измирането на тази голяма и така популярна група животни – динозаврите.
Индикатори за катастрофалните събития са внезапното изчезване на планктона и други морски организми, както и рязката промяна във видовия състав на полена в съответните пластове. Тази промяна свидетелства за внезапно изчезване на покритосеменните растения в края на креда и заместването им от папратови, а след това обратно завръщане на нормалната флора. Този папратов пик е открит в много сухоземни К/Т пластове в западните части на Северна Америка, където фосилните записи от сухоземна растителност са доста подробни и е интерпретиран като следствие на катастрофално количество прах в атмосферата: папратите се възстановяват първи, колонизират освободените площи, последвани от покритосеменните едва след възстановяването на почвите. Това заключение е направено на базата на аналогията с наблюдавани подобни обрати в областта на флората, предизвикани от вулканични изригвания. Разбира се, трябва да бъде отчетен факта, че подобни аномалии са ограничени до фосилните записи на Северна Америка. Навсякъде другаде по света, особено в южното полукълбо, не се наблюдава нищо повече от постепенна промяна във видовия състав на растенията през К/Т периода.
Сред най-популярните заблуди относно К/Т изчезването е тази за степента на влияние на събитията. В действителност повечето групи организми оцеляват. При насекомите, бозайниците, птиците и цветните растения на сушата, както и рибите, коралите и мекотелите в океаните се наблюдава еволюционен разцвет след края на креда. Причината може би се корени в това, че повечето от хипотезите предлагани в миналото, се опитват да обяснят драматичното изчезване само на динозаврите. Не се отчита факта, че К/Т периодът е белязан от глобални събития, така че за обяснението им са нужни причини със съответното въздействие.
За оценка на степента на въздействие и евентуалните промени в екосистемите е необходимо прецизното им и точно реконструиране. Това е предмет на палеоекологията. Вероятно повечето от вас ще бъдат разочаровани да научат, че предмет на палеоекологията не са евентуални палеопротести срещу замърсяващите атмосферата отровни летливи газове на Цехщайново море, нито пък отчаяни палеодействия по съхраняването на българските мезозойски корали, страдащи от необратимите промени в климата. Палеоекологията изучава начина на живот на отделни изкопаеми организми, животни и растения, отношенията помежду им, както и със заобикалящата ги среда, и методите й играят основна роля при проучванията на масовите измирания. Това, разбира се, не винаги е лесно и възможно, като се има предвид непълнотата на фосилните записи и т. нар. ефект на Сигнор-Липс - поради непълни фосилни данни, съществува вероятност повечето измрели видове да са изчезнали реално по-късно от времето, датирано по най-младия намерен фосил.
Масовото измиране в края на креда безспорно е най-популярното от петте големи, главно заради изчезването на енигматичните динозври. До преди 1980 г. относно причините за това са предлагани най-различни хипотези. Те варират от приемливи такива (глобални климатични промени, таксономични промени в растителните съобщества, астероиден сблъсък, тектонични процеси, промени в нивото на световния океан) до по-налудничави (загуба на сексуален апетит, изглупяване и хормонален дисбаланс при динозаврите; ожесточена конкуренция с гъсеници за растителна храна; бозайници, които изяждат всички динозавърски яйца). Множество експедиции и изследвания са проведени с цел да бъде установено какво точно е причинило измирането на тази голяма и така популярна група животни – динозаврите.
През юни 1980 г. в Science е публикувано едно от най-важните изследвания на 20-ти век. В своята публикация Луис Алварес посочва, че 10 километров астероид е ударил Земята, ударът е вдигнал огромно количество прах в атмосферата, покрил е цялото земно кълбо, закривайки слънцето, спира процесът фотосинтеза при сухоземните растения и океанския планктон, което означава изчезване на продуцентите. След изчезването на храната им измират растителноядните животни, последвани от месоядните. Това е в общи линии моделът, описан от Алварес.
 |
| Луис и Уолтър Алварес |
Алварес заключава, че иридиевата аномалия е резултат от сблъсък с огромен метеорит. Той изчислява, че ефектът от подобен сблъсък трябва да се простира по цялото земно кълбо. Въз основа на модели и данни от изригвания на вулкани посочва, че кратерът, оставен от подобен метеорит, трябва да е с размери над 100-150 метра в диаметър, за да произведе подобно количество прах, а за това е необходим метеорит с диаметър приблизително 10 км. Публикацията в Science привлича вниманието на международната научна общност, като коментарите на учени от най-различни области са противоречиви. Отчасти защото авторът е физик, а не палеонтолог или геолог, отчасти защото доказателствата изглеждат недостатъчни. Последвали откритя потвърждават правотата на Алварес и неговия екип, а през 1991 г. бива открит и кратерът – в Мексиканския залив.
За да докаже, че масовото измиране в края на креда е резултат на астероиден сблъсък, Алварес предлага следното просто и дори дръзко уравнение:
M=sA/0.22f
Където М е масата на астероида, s е повърхностното съдържание на иридий след въздействието, А е общата площ на земната повърхност, f е съдържанието на иридий в метеоритите, а 0.22 е количеството материал, изхвърлен от големия вулкан Кракатоа, който изригва през 1883 г., което достига стратосферата. По тази формула Алварес изчислява, че астероидът, причинил К/Т събитията, трябва да е притежавал тегло над 34 милиарда тона. Диаметърът на подобен астероид е определен на 7-10 километра.
Иридиевата аномалия засяга скални пластове от К/Т границата, както на сушата, така и в океана. Подобни пластове са открити на повече от 100 места навсякъде по земното кълбо. Там, където границата е разположена в морски седименти, иридиевият слой е разположен точно над последните кредни микрофосили, а седиментите над него съдържат палеоценски микрофосили от началото на ценозоя. Иридиева аномалия се наблюдава само в скалните проби от К-Т границата и се отличава с много характерен пик, показващ, че събитието е било внезапно и в геологически план краткосрочно.
Освен иридиевата аномалия има и други доказателства в подкрепа на астероидния сблъсък. Известно е, че в пластовете от метеоритни кратери присъстват шоков кварц и микротектити. Микротектитите (земно стъкло) се образуват в резултат на стапяне на скалите от сблъсъка и почти внезапното втвърдяване на материала във въздуха. Шоковият кварц се формира, когато кварцови кристали са подложени на внезапно повишаване на налягането. Това причинява трайни промени под формата на микропукнатини, които лесно се разпознават. Навсякъде в Северна Америка К/Т пластовете са богати на микротектити, а точно над тях е разположен тънък слой, богат на иридий и шоков кварц. Този слой е тънък на места едва няколко милиметра, но общото количество материал надхвърля 1 куб. км. шоков кварц само в западната част на Северна Америка. Зоната на шоковия кварц се разпростира на запад по дъното на Тихия океан, но е рядко срещан в К/Т пластове от останалата част на света. Всичко това показва, че сблъсъкът се е случил някъде в Северна Америка.
Основна алтернатива на астероидния сблъсък като причина за масовото измиране в края на креда е градуалистичният модел, според който изчезванията са се случили в течение на дълги периоди от време, като резултат от периодични климатични промени. На сушата гъстите субтропични гори, обитавани от динозаври, отстъпили място на листопадни гори, доминирани от бозайниците. В подкрепа на градуалистичния модел е плавното понижаване на честотата на срещане на някои групи морски организми в края на креда. Климатичните промени на сушата се свързват с промени в нивото на световния океан и свиване на зоната на топлите плитководни морета.
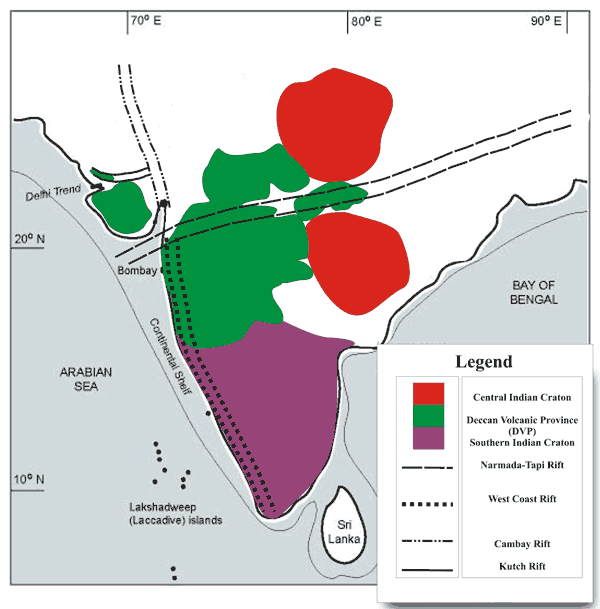
Друга причина, възприемана като основна за масовото К/Т измиране, е вулканична активност. Деканските капани в Индия представляват обширни територии, заливани от лава в продължение на 2-3 милони години през креда. Вулканичният модел обаче не може да даде обяснение на всички налични данни за катастрофата – количеството иридий, шоков кварц, микротектити и др., както и някои биологични последствия.
Откриването на кратера Чиксулуб на полуостров Юкатан, в Централна Америка, окончателно измества градуалистичния и вулканичния модели. Това е яйцевиден по форма кратер, широк около 200 км в диаметър. Скалните пластове под кратера са датирани прецизно към К/Т границата, а скалите около древната брегова линия на протокарибите също подкрепят теорията за сблъсъка - изобилие на шоков кварц. Освен това, в скалите от К/Т границата в протежение на 1000 километра от зоната на сблъсъка се откриват микротектити с уникална геохимия. Микротектитите обикновено се свързват с вулканични изригвания, но тъй като притежават същата геохимия като евапоритите и варовиковите скали, които покриват дъното на кратера, вулканичната хипотеза не може да обясни произхода им.
Яйцевидната форма на кратера Чиксулуб показва, че астероидът е ударил повърхността под малък ъгъл, около 20°–30°, като е разпръснал повече материя на северозапад, отколкото в останалите посоки. Това е основна причина за по-големите поражения на север в американския континент и несиметричното разпределение на шоковия кварц.
Яйцевидната форма на кратера Чиксулуб показва, че астероидът е ударил повърхността под малък ъгъл, около 20°–30°, като е разпръснал повече материя на северозапад, отколкото в останалите посоки. Това е основна причина за по-големите поражения на север в американския континент и несиметричното разпределение на шоковия кварц.
Геоложките и петроложки проучвания, както и иридиевата аномалия, шоковият кварц и микротектитите, а и кратерът Чиксулуб, дават силни доказателства в подкрепа на астероиден удар преди 65 млн. години. Данните от палеонтологията обаче говорят за внезапно изчезване на някои групи организми, докато други се характеризират с по-продължително такова, за периоди от около 1-2 млн. години. Основният въпрос е как точно сблъсъкът е причинил изчезването. Наличните модели за измирането са биологически малко вероятни или твърде катастрофични: сценарият трябва да отчита факта, че 75% от семействата преживяват К/Т събитията, повечето от които почти не биват засегнати. Дали двата модела могат да бъдат комбинирани така, че дългосрочните промени да бъдат обяснени чрез постепенна промяна в океанското ниво и климата, а финалното измиране в края на креда да е резултат на сблъсъка, е трудно да се каже.
Няма никакво съмнение, че северноамериканският континент е бил напълно опустошен. В световен мащаб дори краткосрочни екстремни събития, повлияващи сухоземната растителност и морския планктон, биха причинили срив в хранителните вериги. Основните глобални ефекти от астероидния сблъсък според повечето модели се изразяват в намаляване на осветеността, резки температурни промени, киселинни дъждове и пожари. Облакът от прахови частици в атмосферата се простира по цялото земно кълбо, като силно намалява осветеността и инхибира фотосинтезата. Теоретични и експериментални данни потвърждават, че значителна промяна в осветеността в продължение на няколко дни може да унищожи до 99% от планктона. Вероятността за това се увеличава при положение, че са замесени и обширни пожари, тъй като саждите абсорбират слънчевата радиация по-ефективно от праховите частици.
Логично е птерозаврите, динозаврите и големите морски влечуги да са уязвими по отношение на хранителен недостиг и подобно обяснение за тяхното измиране - с изчезване на продуцентите в хранителните вериги - изглежда правдоподобно. От друга страна оцелелите гущери и примитивни бозайници са малки по размери животни, често ровещи или хиберниращи, способни да си набавят ядки, семена, ларви и други безгръбначни за храна. В океаните по-уязвими са безгръбначните обитаващи плитководната зона, където температурните промени могат да бъдат резки и фатални. За разлика от дълбоководните обитатели, които не са подложени на температурен шок, характеризират се с много по-нисък метаболизъм и са способни да издържат на няколко месеца гладуване. Фауната в големите географски ширини е адаптирана към периоди на зимен мрак, затова тропическите съобщества са по-уязвими. Листопадните дървета преживяват К/Т събитията много по-успешно от вечнозелените. Т.е. видовете обитаващи големите географски ширини и дълбоките океански води са имали по-добри шансове. Също така сладководните съобщества са имали предимство пред сухоземните такива. Фосилни находки от Северна Дакота показват, че костенурките и други водни влечуги преминават успешно К/Т границата, докато динозаврите изчезват напълно. Като се има предвид, че сладководните съобщества се поддържат до голяма степен от потока детрит, включващ предимно сухоземна растителност, лесно е да се предположи, че видовете в хранителните вериги, които се изхранват с детрит, ще преживеят катастрофата по-успешно от останалите.
Единственото пълно и сравнително богато на фосили разкритие на сухоземни пластове от К/Т периода се намира в северноамериканския запад, източна Монтана. Анализът на фосилните находки не показва спад в числеността и разнообразието на динозаврите до края на креда, когато те изчезват внезапно от записите. Това, разбира се, не слага край на спора между катастрофисти и градуалисти. Фосилни находки от други региони свидетелстват за постепенен спад в разнообразието от края на кампан, характеризиращ се с 45 рода, до късен маастрихт (последният етаж на креда) - 24 рода. Това е силно доказателство в подкрепа на твърдението, че динозаврите вече са преживявали своя залез, а астероидният сблъсък просто е изиграл ролята на coup de grâce.
Останалите групи гръбначни животни - сухоземни и морски - също не се отличават с кой знае колко пълни фосилни записи. Вижда се, че на сушата двуутробните бозайници са по-сериозно засегнати от плацентните. Но тново има много малко доказателствен материал извън западните части на Северна Америка. Крокодилите и сладководните костенурки преживяват К/Т събитията почти незасегнати. Архаичните видове птици изглежда изчезват, а по-съвременните такива се характеризират с широко разпространение в началото на терциер, но фосилните записи са доста непълни, а между позициите на палеонтолозите и молекулярните биолози има несъгласие по този въпрос: последните твърдят, въз основа на филогенетичните си реконструкции, че произходът на много съвременни разреди птици може да бъде проследен назад до креда.
Друга голяма група гръбначни животни са птерозаврите, които без съмнение са летели както над сушата, така и над моретата, но техни добри фосилни останки се намират единствено в морски седименти. Птерозаврите изчезват в края на креда, както и ихтиозаврите и плезиозаврите. Но отново фосилните записи не са достатъчно подробни, за да се твърди със сигурност как точно е протекло изчезването им. Единствено при ихтиозаврите и плезиозаврите има достатъчно данни за продължителен, плавен спад в числеността и разнообразието преди края на креда. Категорична подкрепа от страна на гръбначните животни в полза на катастрофичния модел предлагат гигантските морски влечуги - мозазаврите - които процъфтяват през креда, само за да изчезнат драматично в края на периода.
Най-пълни са фосилните записи на планктонните организми - коколити и фораминифери. В морските седименти, в които тези микрофосили присъстват в изобилие, се наблюдава рязък видов обрат за много кратък период от време в края на креда. Разбира се, съществуват противоречиви становища и по този въпрос. И докато коколитите подкрепят катастрофичния сценарий, при фораминиферите е доста по-неясно как точно са протекли събитията, като според някои палеонтолози има видове, преминали К/Т границата. Но това, освен от подробните фосилни данни, зависи до голяма степен и от качественото им таксономично определяне.
Друга голяма група сухоземни животни, характеризираща се с богати фосилни записи, са насекомите. За разлика от масовото измиране в края на перм, когато насекомите биват сериозно засегнати, през К/Т периода не се наблюдава особена промяна в техния видов състав.
Някои важни мезозойски групи животни, като амонитите, са в залеза си в края на креда, докато при други видове от същите пластове се наблюдава рязко изчезване от фосилните записи. Освен бивалвиите, други две важни групи мекотели, рудисти и иноцерамиди - и двете характерни за периода креда - изчезват вероятно 1 млн. години преди края на мастрихт.
Друг основен проблем, с който повечето катастрофични сценарии не могат да се справят, е определянето на пола от температурата на околната среда при някои (ектотермни) видове. Това е характерно за много съвременни влечуги. Полът при тези видове не се определя генетично, а от температурата на околната среда по време на ембрионалното развитие. Най-често при по-високи температури се развива полът, който се характеризира с по-големи размери във възрастно състояние. Женските костенурки, например, са по-едри от мъжките, защото произвеждат голямо количество яйца. Съответно ембрионите се развиват като женски възрастни индивиди, ако температурата на средата е по-висока. При крокодилите и гущерите е обратно. Мъжките са по-едри от женските, защото при тях се наблюдава много силно съперничество. Определянето на пола от околната среда е много разпространено сред съвременните влечуги и вероятно е съществувало при техните предшественици. При това положение една внезапна промяна в условията на околната среда би била фатална за повечето влечуги през К/Т периода. Нещо, което не се наблюдава в пластовете.
Оцеляването на птиците също е своеобразна загадка. Дребните видове динозаври не се отличават от по-едрите видове птици, както по размери, така и в екологично отношение. Птиците обаче оцеляват, а динозаврите не. Птиците са топлокръвни животни с висок метаболизъм, много чувствителни на екстремни промени. При все това те оцеляват. Предполага се, че това се дължи на способностите им да се гмуркат, плуват или търсят убежища във водата, в блата и мочурливи зони, да си правят дупки и да гнездят в хралупи на дървета, като по този начин се укриват от непосредственото влияние на околната среда.
В обобщение може да се каже, че нито катастрофичните, нито градуалистичните модели могат да предложат самостоятелно обяснение на наличните данни за събитията от края на креда и изводите направени от тях в световен мащаб.
Ясно е, че екстремните катастрофични модели трябва да бъдат преразгледани, за да отговарят на степента на изчезване. Много групи преживяват събитията, почти няма големи таксони, които да са напълно заличени. Дори динозврите оцеляват в известен смисъл - като птици. Все още не са напълно ясни причините за изчезването на толкова много групи организми в края на креда, докато други оцеляват. Не е ясно дали това е причинено само от астероидния сблъсък или от комбинацията на сблъсъка с вулканизъм, както и последователността на тези явления, и на биологичните и екологични последствия от тях. Необичайният екстремен ефект от К/Т събитията, глобалният обхват и екологичните аномалии (като папратовия скок) може би се дължат на това, че астероидното въздействие и вулканизмът са се случили в период, в който равновесието в глобалните екосистеми вече е било твърде уязвимо.
Няма никакво съмнение, че северноамериканският континент е бил напълно опустошен. В световен мащаб дори краткосрочни екстремни събития, повлияващи сухоземната растителност и морския планктон, биха причинили срив в хранителните вериги. Основните глобални ефекти от астероидния сблъсък според повечето модели се изразяват в намаляване на осветеността, резки температурни промени, киселинни дъждове и пожари. Облакът от прахови частици в атмосферата се простира по цялото земно кълбо, като силно намалява осветеността и инхибира фотосинтезата. Теоретични и експериментални данни потвърждават, че значителна промяна в осветеността в продължение на няколко дни може да унищожи до 99% от планктона. Вероятността за това се увеличава при положение, че са замесени и обширни пожари, тъй като саждите абсорбират слънчевата радиация по-ефективно от праховите частици.
Логично е птерозаврите, динозаврите и големите морски влечуги да са уязвими по отношение на хранителен недостиг и подобно обяснение за тяхното измиране - с изчезване на продуцентите в хранителните вериги - изглежда правдоподобно. От друга страна оцелелите гущери и примитивни бозайници са малки по размери животни, често ровещи или хиберниращи, способни да си набавят ядки, семена, ларви и други безгръбначни за храна. В океаните по-уязвими са безгръбначните обитаващи плитководната зона, където температурните промени могат да бъдат резки и фатални. За разлика от дълбоководните обитатели, които не са подложени на температурен шок, характеризират се с много по-нисък метаболизъм и са способни да издържат на няколко месеца гладуване. Фауната в големите географски ширини е адаптирана към периоди на зимен мрак, затова тропическите съобщества са по-уязвими. Листопадните дървета преживяват К/Т събитията много по-успешно от вечнозелените. Т.е. видовете обитаващи големите географски ширини и дълбоките океански води са имали по-добри шансове. Също така сладководните съобщества са имали предимство пред сухоземните такива. Фосилни находки от Северна Дакота показват, че костенурките и други водни влечуги преминават успешно К/Т границата, докато динозаврите изчезват напълно. Като се има предвид, че сладководните съобщества се поддържат до голяма степен от потока детрит, включващ предимно сухоземна растителност, лесно е да се предположи, че видовете в хранителните вериги, които се изхранват с детрит, ще преживеят катастрофата по-успешно от останалите.
Единственото пълно и сравнително богато на фосили разкритие на сухоземни пластове от К/Т периода се намира в северноамериканския запад, източна Монтана. Анализът на фосилните находки не показва спад в числеността и разнообразието на динозаврите до края на креда, когато те изчезват внезапно от записите. Това, разбира се, не слага край на спора между катастрофисти и градуалисти. Фосилни находки от други региони свидетелстват за постепенен спад в разнообразието от края на кампан, характеризиращ се с 45 рода, до късен маастрихт (последният етаж на креда) - 24 рода. Това е силно доказателство в подкрепа на твърдението, че динозаврите вече са преживявали своя залез, а астероидният сблъсък просто е изиграл ролята на coup de grâce.
Останалите групи гръбначни животни - сухоземни и морски - също не се отличават с кой знае колко пълни фосилни записи. Вижда се, че на сушата двуутробните бозайници са по-сериозно засегнати от плацентните. Но тново има много малко доказателствен материал извън западните части на Северна Америка. Крокодилите и сладководните костенурки преживяват К/Т събитията почти незасегнати. Архаичните видове птици изглежда изчезват, а по-съвременните такива се характеризират с широко разпространение в началото на терциер, но фосилните записи са доста непълни, а между позициите на палеонтолозите и молекулярните биолози има несъгласие по този въпрос: последните твърдят, въз основа на филогенетичните си реконструкции, че произходът на много съвременни разреди птици може да бъде проследен назад до креда.
Друга голяма група гръбначни животни са птерозаврите, които без съмнение са летели както над сушата, така и над моретата, но техни добри фосилни останки се намират единствено в морски седименти. Птерозаврите изчезват в края на креда, както и ихтиозаврите и плезиозаврите. Но отново фосилните записи не са достатъчно подробни, за да се твърди със сигурност как точно е протекло изчезването им. Единствено при ихтиозаврите и плезиозаврите има достатъчно данни за продължителен, плавен спад в числеността и разнообразието преди края на креда. Категорична подкрепа от страна на гръбначните животни в полза на катастрофичния модел предлагат гигантските морски влечуги - мозазаврите - които процъфтяват през креда, само за да изчезнат драматично в края на периода.
Най-пълни са фосилните записи на планктонните организми - коколити и фораминифери. В морските седименти, в които тези микрофосили присъстват в изобилие, се наблюдава рязък видов обрат за много кратък период от време в края на креда. Разбира се, съществуват противоречиви становища и по този въпрос. И докато коколитите подкрепят катастрофичния сценарий, при фораминиферите е доста по-неясно как точно са протекли събитията, като според някои палеонтолози има видове, преминали К/Т границата. Но това, освен от подробните фосилни данни, зависи до голяма степен и от качественото им таксономично определяне.
Друга голяма група сухоземни животни, характеризираща се с богати фосилни записи, са насекомите. За разлика от масовото измиране в края на перм, когато насекомите биват сериозно засегнати, през К/Т периода не се наблюдава особена промяна в техния видов състав.
Някои важни мезозойски групи животни, като амонитите, са в залеза си в края на креда, докато при други видове от същите пластове се наблюдава рязко изчезване от фосилните записи. Освен бивалвиите, други две важни групи мекотели, рудисти и иноцерамиди - и двете характерни за периода креда - изчезват вероятно 1 млн. години преди края на мастрихт.
Друг основен проблем, с който повечето катастрофични сценарии не могат да се справят, е определянето на пола от температурата на околната среда при някои (ектотермни) видове. Това е характерно за много съвременни влечуги. Полът при тези видове не се определя генетично, а от температурата на околната среда по време на ембрионалното развитие. Най-често при по-високи температури се развива полът, който се характеризира с по-големи размери във възрастно състояние. Женските костенурки, например, са по-едри от мъжките, защото произвеждат голямо количество яйца. Съответно ембрионите се развиват като женски възрастни индивиди, ако температурата на средата е по-висока. При крокодилите и гущерите е обратно. Мъжките са по-едри от женските, защото при тях се наблюдава много силно съперничество. Определянето на пола от околната среда е много разпространено сред съвременните влечуги и вероятно е съществувало при техните предшественици. При това положение една внезапна промяна в условията на околната среда би била фатална за повечето влечуги през К/Т периода. Нещо, което не се наблюдава в пластовете.
Оцеляването на птиците също е своеобразна загадка. Дребните видове динозаври не се отличават от по-едрите видове птици, както по размери, така и в екологично отношение. Птиците обаче оцеляват, а динозаврите не. Птиците са топлокръвни животни с висок метаболизъм, много чувствителни на екстремни промени. При все това те оцеляват. Предполага се, че това се дължи на способностите им да се гмуркат, плуват или търсят убежища във водата, в блата и мочурливи зони, да си правят дупки и да гнездят в хралупи на дървета, като по този начин се укриват от непосредственото влияние на околната среда.
В обобщение може да се каже, че нито катастрофичните, нито градуалистичните модели могат да предложат самостоятелно обяснение на наличните данни за събитията от края на креда и изводите направени от тях в световен мащаб.
Ясно е, че екстремните катастрофични модели трябва да бъдат преразгледани, за да отговарят на степента на изчезване. Много групи преживяват събитията, почти няма големи таксони, които да са напълно заличени. Дори динозврите оцеляват в известен смисъл - като птици. Все още не са напълно ясни причините за изчезването на толкова много групи организми в края на креда, докато други оцеляват. Не е ясно дали това е причинено само от астероидния сблъсък или от комбинацията на сблъсъка с вулканизъм, както и последователността на тези явления, и на биологичните и екологични последствия от тях. Необичайният екстремен ефект от К/Т събитията, глобалният обхват и екологичните аномалии (като папратовия скок) може би се дължат на това, че астероидното въздействие и вулканизмът са се случили в период, в който равновесието в глобалните екосистеми вече е било твърде уязвимо.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}