Приматоподобни бозайници.
През Палеоцен повечето приматоподобни бозайници са принадлежали към клас Plesiadapiformes. Плезиадапиформите са считани за архаични членове на Primates. Въпреки че плезиадапиформите имат сходства със съвременните примати по някои характеристики на скелета, те са били на много по-ниско еволюционно стъпало, подобно на днешните дървесни земеровки. Съвременните примати са несравними сред бозайниците по отношение на адаптациите си за живот по дърветата. Техните способности за захващане и скачане им позволяват бързо придвижване в тази среда, което е свързано и с развитието на по-голям мозък. Доколкото е известно, плезиадапиформите също са прекарвали живота си по дърветата. Въпреки това, при тях липсват адаптации за бързо скачане, каквито наблюдаваме при днешните примати и не са били способни на толкова бързо придвижване сред дърветата. Освен това мозъкът им е бил доста по-малък в сравнение с днешните примати. От друга страна, плезиадапиформите развиват някои характеристики, които не се наблюдават при по-късните примати – например уголемените резци, сходни с тези на гризачите. Това показва, че плезиадапиформите не са директни предшественици на приматите, а по-скоро представляват разклонение, което се отделя в ранен стадий от основния клон. Картината се усложнява от съществуващите и днес разреди бозайници, които би трябвало да бъдат причислени към родословното дърво на приматите – дървесните земеровки от разред Scandentia, летящите лемури от разред Dermoptera, а може би и прилепите (Chiroptera), въпреки че според последните молекулярни изследвания те са далечни роднини. Къде точно сред тях трябва да бъдат вместени плезиадапиформите е все още спорен въпрос. Много учени считат за погрешно причисляването им към приматите и ги отделят в нов разред – Plesiadapiformes.
 |
Реставрация на Purgatorius, един от най-ранните известни приматоподобни бозайници. |
Най-примитивният известен вид сред плезиадапифомите е Purgatorius, животно с размерите на плъх. Намерени са останки само от зъби и челюсти. Тяхното устройство отговаря на модела, характерен за общите предшественици на всички плацентни бозайници – насекомоядните. Зъбите на Purgatorius имат черти, характерни както за плезиадапиформите, така и за истинските примати. Тяхната форма предполага, че животните са се хранили не само с насекоми, но и с плодове. В реставрациите Purgatorius често са представяни като бягащи от къснокредни динозаври като Tyrannosaurus или Triceratops. Всъщност почти всички фосилни находки на Purgatorius са от раннопалеоценски пластове в Северна Америка. Един единствен зъб е открит заедно с останки на динозаври и този зъб най-вероятно е „замърсяване” от палеоценски скали, които вероятно са били смесени с кредни по погрешка, когато палеонтолозите са пресявали останките. Друг възможен сценарий е депозитите, съдържащи зъбът на пургаториус да са били отложени в началото на Палеоцен, като реката ги врязва дълбоко в кредните пластове, смесвайки палеоценските останки на бозайниците с кредните останки от динозаври. Дори и да са съществували приматоподобни бозайници през Мезозой, доказателства за това все още няма. Ако искаме да разберем повече за ранните приматоподобни бозайници, отколкото ни позволяват намерените зъби и челюсти, трябва да погледнем към по-късните, специализирани форми. Най-добре изучените плезиадапиформи са тези от сем. Plesiadapidae – едно от най-преуспелите семейства палеоценски бозайници. Плезиадапидите са били животни с размери на катерица или плъх, притежаващи зъбна редица, подобна на тази у гризачите, с двойка здрави, уголемени резци, които у по-късните форми са последвани от диастема. Докато долните резци са били с по-просто устройство, формиращи нещо като гребло заедно, горните са били много по-сложно устроени, с три отделни гънки отпред и една отзад. За разлика от резците при гризачите, тези на плезиадапидите не са притежавали самозаточващ ръб, нито са имали неограничен растеж, затова вероятно не са използвани за дълбаене. При по-съвременните плезиадапиди дъвкателните зъби са по-плоски, а емайлът им по-назъбен (грапав), което показва, че са приемали предимно растителна храна – вероятно листа или плодове.
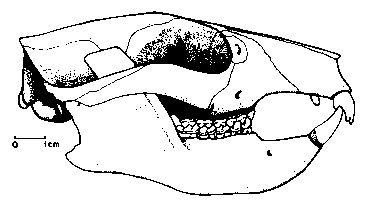 |
Череп на Plesiadapis tricuspidens. |
По-пълни черепни и скелетни находки има от къснопалеоценският род Plesiadapis. Този род вероятно възниква в Северна Америка и се разпространява в Европа по суша през днешна Гренландия. Благодарение на високата численост на Plesiadapis и неговата бърза еволюция, видовете му играят основна роля при разпределението и датирането на къснопалеоценските континентални седименти и връзките между фауната от двете страни на Атлантика. Две изключителни скелетни находки на плезиадаписи (единият почти пълен) бяха открити в езерни седименти в Менат, Франция. Въпреки че находките са лошо съхранени, има следи от кожа и косми по тях – нещо уникално за палеоценски бозайници. Други скелетни находки на плезиадаписи показват, че животните са били често срещани в този регион. Черепът е сходен с тези на гризачите, широк и плосък с остра муцуна и все още странично разположени очни орбити, за разлика от разположените отпред очи на днешните примати, подпомагащи триизмерното виждане. Черепната кухина е по-малка в сравнение с днешните стандарти, но по-голяма от тази на живеещите по това време копитни бозайници, например. Плезиадаписите са имали много подвижни крайници, завършващи със силно извити нокти и дълга, пухкава опашка, която е чудесно съхранена при скелетите от Менат. Начинът на живот на плезиадаписите е много обсъждан в миналото. Като родствени на приматите те вероятно са се катерили по дърветата, но останките на дървесни форми са изключително редки. Затова повечето учени приемат, че плезиадаписите са живеели на земята, подобно на днешните мармоти и земни катерици. По-съвременни проучвания показват, че плезиадаписите са били умели катерачи, подобно на дървесните катерици и опосумите. 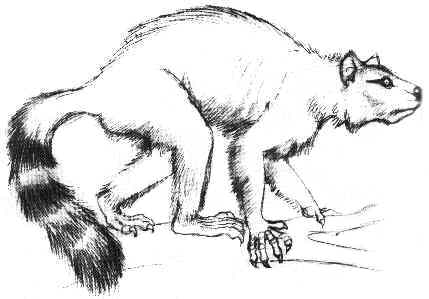 |
Реставрация на Plesiadapis. |
Второто семейство на плезиадапиформите, Carpolestidae, е много характерно за Северна Америка, въпреки че никога не доминира така, както плезиадапидите. Карполестидите също притежават уголемени резци. Долните са по-малки от горните, които притежават характерна вдлъбнатина, в която се поместват точно ръбовете на долните резци. Карполестидите са много по-дребни, с размерите на мишка или плъх. Последните долни предкътници при карполестидите развиван изключително остри, големи и назъбени гънки. Наречени са плагиаулакоиди и се развиват независимо в няколко групи бозайници, най-изразени са при мултитуберкулатите. При карполестидите тези зъби се противопоставят на също уголемените, но по-плоски трети и четвърти предкътник на горната челюст. Този дъвкателен механизъм се развива за обработка на растителна храна, съдържаща много фибри като плодове, семена, а вероятно и малки насекоми. Развитието на този високоспециализирана адаптация може да бъде проследена при северноамериканските родове Elphidotarsius, Carpodaptes и Carpolestes, които оформят основната линия в еволюцията на карполестидите. Дребничките, с размерите на мишка, Elphidotarsius от средния Палеоцен на Северна Америка са подобни на примитивните плезиадапиди, но последните им долни предкътници вече напомнят копиеподобната форма на тези при по-съвременните карполестиди. В късен Палеоцен основните черти на семейството са добре представени у Carpodaptes и се утвърждават и усъвършенстват при къснопалеоценските видове на Carpolestes, последните северноамерикански карполестиди.
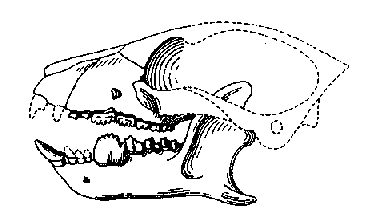 |
Череп на къснопалеоценски Carpodaptes. |
 |
Реставрация на Carpolestes simpsoni. |
Друго семейство високоспециализирани плезиадапиформи са Picrodontidae, дребни животинки представени от родът Picrodus, който присъства само с няколко вида през среден и късен Палеоцен в Северна Америка. Пикродонтидите притежават дълги, ножоподобни долни резци, подобно на останалите плезиадапиформи, но дъвкателните им зъби са силно модифицирани. Първите горен и долен кътник са силно уголемени, а всички останали кътници формират широки, плоски повърхности със силно нагънат емайл. Наблюдава се увеличаване на общата площ, което е необходимо при смилането на някаква мека храна. Така специализираните кътници са толкова сходни с тези на съвременните прилепи, че някои изследователи причисляват пикродонтидите към същия разред. Днес тази прилика се счита за резултат от конвергенция, но свидетелства за хранителните навици на пикродонтидите. Също както и тези прилепи, пикродонтидите вероятно са се хранили с цветен нектар и полен, висококалорична храна, която не натоварва зъбите, както и дървесни сокове и сочни плодове.
По-дребните видове плезиадапиформи принадлежат към семейство Micromomyidae. Семейството включва миниатюрни форми от късен Палеоцен до ранен Еоцен в региона на Скалистите Планини, които носят подходящи имена като Micromomys и Tinimomys (наставката mys означава мишка). Тези животни са достигали тегло около 30 грама – много по-малко в сравнение с най-дребния съвременен примат, мишият лемур Microcebus от Мадагаскар. Цялостен скелет на все още неименуван вид микромомид бе открит в къснопалеоценски утаечни скали. Той принадлежи на дребно животинче, тежащо едва 20 грама, което е било високоспециализирано за катерене по дърветата и вероятно не е слизало на земята. Бозайници с подобен размер на тялото са предимно насекомоядни и всъщност острите зъби на Micromomys потвърждават, че са ловували предимно насекоми.
Други плезиадапиформи вероятно са събирали плодове сред широкоразпространените по това време тропически и субтропически гори. Членовете на Paromomyidae се характеризират с плоски, квадратни кътници, формиращи широка повърхност, подходяща за смилане на плодове. Дървесни сокове и дребни насекоми вероятно са допълвали диетата им. Зъбните характеристики на паромомидите могат да бъдат проследени назад до Paromomys от среден Палеоцен на Северна Америка. При по-развитите форми като Ignacius и Phenacolemur са силно удължени и тънки, а след тях се развива голяма диастима, както при плезиадапидите. И двата рода са се срещали от късен Палеоцен до среден Еоцен. По този начин те се оказват последните оцелели плезиадапиформи – живи фосили в свят, вече населен от истинските примати.
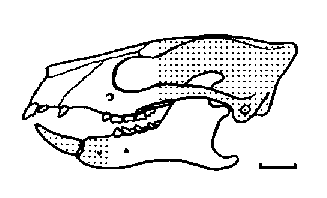 |
Череп на Ignacius graybullianus. |
Въпреки че няма доказателства за близкородствени отношения между плезиадапиформите и летящите лемури, друга група палеоценски бозайници вероятно са първите дермоптери. Сред тях са плагиоменидите, странна група, която се появява първо през среден Палеоцен в Северна Америка. Плагиоменидите са рядко срещани представители на палеоценската фауна, но те могат да доминират в отделни местообитания, което показва високоспециализирани изисквания. Фосили на къснопалеоценския плагиоменид Planetetherium, например, са често срещани във въгледобивни мини в Монтана, но не се срещат почти никъде другаде. Животните вероятно са намерили идеални условия за живот в гъстите, заблатени гори в тази местност. През Еоцен някои плагиомениди дори процъфтяват на остров Елсмир, най-северната точка на Канада, която се пресича от арктичната окръжност. Климатът тогава е бил много по-топлъл от днешния, разбира се, но на такава географска ширина (72 градуса север, по това време) животните вероятно са били изправени пред по-дългите зимни нощи.
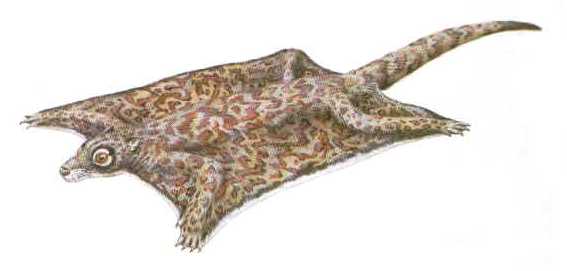 |
Хипотетична реставрация на Planetetherium. |
Плезиадапиформите и плагиоменидите са сред първите приматоподобни бозайници, които процъфтяват през Палеоцен. Тези архаични форми биват заменени през Еоцен от истинските примати (еурипримати). Най-ранните известни еурипримати принадлежат към семействата Adapidae и Omomyidae. И двете групи притежават адаптации за захващане и скачане, както и сравнително голям мозък, подобен на днешните примати. Адапидите са подобни, а вероятно и родствени на лемурите в Мадагаскар, както и на лоритата от тропическите части на Африка и Азия. Те включват главно едри животни, активни предимно денем. Типичните омомиди са дребни животни с огромни очи, които свидетелстват за нощна активност. Омомидите вероятно включват предшествениците на днешните дългопети от Югоизточна Азия, нощни хищници на дребна плячка, които са много добри катерачи и се придържат към клоните във вертикална позиция. Адапидите и омомидите се появяват внезапно във фосилните находки на Европа и Северна Америка от началото на еоцен, което показва, че те са имигрирали по това време там от друга част на света, вероятно от Африка или Азия. Предполагаеми еурипримати са описани от палеоцена на тези континенти. Petrolemur и Decoredon от палеоцена на Китай са описани като ранни представители на адапидите и омомидите респективно. Те обаче са слабо познати и не е доказано, че принадлежат към приматите въобще. По-обещаващ кандидат е Altiatlasius от късния палеоцен на Мароко, едно от малкото известни ни животни от палеоцена на Африка. Първоначално определен като омомид, Altiatlasius изглежда потвърждава ролята на Африка като център на еволюцията на приматите. Все пак скорошно проучване стигна до извода, че Altiatlasius не са еурипримати, а принадлежат към предимно европейската група плезиадапиформи, които се разпространяват в Африка през Тетис.
Така че все още са непълни доказателствата за ранната история на истинските примати – история, която започва без съмнение през палеоцен, ако не и по-рано. Въпреки това имаме достатъчно доказателства за невероятния успех, който постигат техните родственици – плезиадапиформите – в началото на неозоя. Както видяхме, зъбите на плезиадапиформите свидетелстват за разнообразие от хранителни стратегии – от насекомоядни до пълни вегетарианци и хранещи се нектар и полен видове и тепърва предстои да изясним и други аспекти от биологията им, като придвижването.
Няма коментари:
Публикуване на коментар