Насекомоядни бозайници
Традиционно насекомоядните са класифицирани като клон на плацентните бозайници – Insectivora. Групата включва познати животни като земеровка, таралеж, къртица, както и по-екзотичните форми като златните къртици от Африка, тенрекът от Мадагаскар. Както показва името, тези животни са се хранили с малки безгръбначни като насекоми или червеи, но всъщност менюто на повечето насекомоядни е включвало почти всичко органично, което може да бъде сдъвкано. Въпреки че повечето насекомоядни са развили функционални адаптации, характеристики като много малкия мозък показват, че те са сред най-примитивните плацентни бозайници. Ранните изследвания показват, че насекомоядните заемат централно място сред плацентните бозайници и представляват основно звено, от което еволюират всички останали плацентни. Новооткрити фосили на примитивни форми, които не показват очевидна връзка с някоя от по-развитите групи, биват включвани към насекомоядните по подразбиране. По този начин разредът Насекомоядни се превръща в „кошче за боклук” за проблематични, предимно примитивни плацентни бозайници. Тази традиционна концепция за насекомоядните, която приема групата като основа за развитието на плацентните бозайници днес е отхвърлена. Вместо това се предполага, че днешните насекомоядни и техните близки изкопаеми роднини са отделен клон в еволюционното дърво на плацентните бозайници, също както гризачите или приматите, и не трябва да се свързват с неродствените архаични бозайници. За да се избегне объркването със старата концепция за насекомоядните, често се използват различни наименования – Lipotyphla – за разреда, включващ „истинските” насекомоядни, а за архаичните форми – Proteutheria. За да направят нещата още по-сложни, молекулярни изследвания показват, че някои насекомоядни от Африка и Мадагаскар - златните къртици и тенреците - не са близкородствени на „по-северните” насекомоядни. Вместо това те са част от типично африканско разклонение на бозайниците – Afrotheria, към което принадлежат и слоновете, морските крави, даманите и тръбозъбите. Поради това златните къртици и тенреците бяха преместени от Lipotyphla в нов разред – Afrosoricida. За удобство терминът „насекомоядни” ще се използва тук за животните подобни на насекомоядни, независимо дали те принадлежат към Lipotyphla или Afrosoricida.
 |
| Cimolestes |
Отдавна е известно, че насекомоядните бозайници са съществували през късна Креда. Cimolestes, например, процъфтяват в края на Креда в Северна Америка и този род дори оцелява до началото на Палеоцен. Повечето видове Cimolestes са били с размерите на плъх или заек, но в края на Креда Cimolestes magnus достига размерите на мармот, което го прави един от най-големите известни мезозойски бозайници. Въпреки че класифицирането на толкова ранни форми все още подлежи на обсъждане, те често се причисляват към сем. Palaeoryctidae. Основните членове на групата се появяват в началото на Терциер и в Северна Америка включват Palaeoryctes от средата до края на Палеоцен. Повечето видове на Palaeoryctes варират от 20г до 60г на тегло, което ги поставя по размери между мишка и катерица. Кътниците им са притежавали високи и остри ръбове. Те изглежда са изпълнявали предимно пробивна функция, необходима за проникване през твърдите външни скелети на насекомите. Когато Palaeoryctes са били описани за първи път, е било на базата на само един скелет, затова името им (означаващо „ранни ровещи”) се базира на предположения. Когато по-късно бива открит фрагмент от хумеруса на животно, става ясно, че Palaeoryctes са били добре приспособени за копаене и името им става повече от подходящо.
 |
| Palaeoryctes |
Видове като палеориктидите показват, че разликата между насекомоядните и дребните месоядни бозайници е малка. Зъбната редица на Cimolestes, например, хвърля светлина върху режещите структури, наблюдавани при месоядните. Докато малките видове изглежда са били типичните насекомоядни, по-големите Cimolestes magnus вероятно са ловували по-едра плячка и в известна степен са били хищници. Всъщност е възможно двете големи групи плацентни хищници, разредът Creodonta и истинските хищници от разред Carnivora, да са еволюирали от подобни на Cimolestes видове. Вероятните родственици на палеориктидите – видовете Hyracolestes ermineus и Sarcodon pygmaeus са били често срещани през късен Палеоцен в Монголия и Китай и са заемали нишата на дребните хищници там. Hyracolestes са тежали около 40 грама, а Sarcodon – 75г., което е приблизително колкото днешните невестулки. Като белег за свирепите навици на своите собственици, дъвкателните зъби на Hyracolestes показват характеристики, сходни с тези на съвременните хищници – способност да задържат храната неподвижна, докато бъде срязвана от острите ръбове.
Палеориктидите може би са еволюирали и в малко по-различна посока, зъбите на някои от тях много напомнят тези на архаичинте бозайници като Taeniodonta – сравнително едри животни, специализирани за копаене. Палеориктидът Alveugena от Уайоминг дори е бил смятан за междинно звено между двете групи. Това са най-едрите известни палеориктиди, достигащи размерите на куче. Но откритието на мезозойския тениодонт Schowalteria (от късна Креда) в Албърта отново възроди споровете относно тази хипотеза. Освен факта, че Schowalteria е много по-стар като вид от „междинното звено” Alveugen, зъбите на тениодонтите са много по-примитивни в повечето отношения, което предполага, че много (дори всички) прилики между ранните тениодонти и палеориктидите се дължат на паралелна еволюция.
Някои учени смятат, че палеориктидите са членове на Lipotyphla (истинските насекомоядни), но това е все още спорен въпрос. Къснокредните Paranyctoides са смятани за най-старите известни Lipotyphla и нищо не подкрепя мезозойския произход на разреда, но фактите са все още оскъдни. Първите безспорни Lipotyphla са известни от Палеоцен и те са включвали видове от двете големи групи съвременни липотифла – таралежите от една страна, и земеровките и родствени на тях видове като къртиците от друга. Членовете на двете групи са сред най-дребните бозайници запазени от палеоценската фауна.
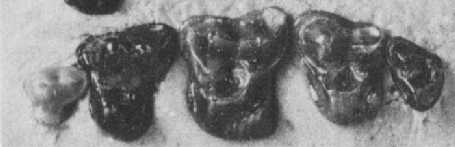 |
Горни дъвкателни зъби на Adapisorex abundans |
Mackennatherium от среден Палеоцен е сред първите предшественици на таралежите в Северна Америка, познати като еринацеоморфи (от Erinaceus, европейския таралеж), както и близкородственият Adunator от Германия. Това са били животни с размерите на мишка, тежащи около 10-20 грама, които имат по-малки гънки на дъвкателните си зъби в сравнение с палеориктидите или останалите насекомоядни. Зъбите им много напомнят тези на копитните бозайници от сем. Hypsodontidae, а класифицирането на няколко вида е все още въпрос на спорове. Няколко къснопалеоценски насекомоядни - като Litolestes от Уайоминг и Adapisorex от Франция и Германия – имат много сходства с таралежите, затова са класифицирани в същото семейство Erinaceidae. Това всъщност е изключение, защото всички останали плацентни бозайници от Палеоцен, с изключение на броненосците, принадлежат към семейства, които днес не съществуват. Класифицирането им като членове на семейство Erinaceidae не означава, че тези ранни насекомоядни са притежавали защитните механизми на таралежите. Дори не всички днешни представители на това семейство притежават бодли. Както показва един изключително добре запазен фосил от Германия, у таралежоподобния Macrocranion се развиват бодли в средата на Еоцен. Родът Macrocranion, познат още и от къснопалеоценски фосилни находки в Уайоминг, е притежавал удължени задни крайници и дълга опашка. Очевидно животните са можели да подскачат на задните си крака (вероятно за избягване на хищници), което е рядко срещана форма на движение при таралежоподбни. |
| Macrocranion tupaiodon |
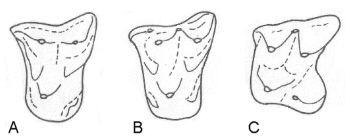 |
| Еволюция на диламбдодонтизма: Leptacodon(A), Pontifactor(B), Sylvisorex(C). |
 |
| Почти пълна долна челюст на Ceutholestes dolosus. |
Поради недостига на африкански фосили на бозайници от ранен Терциер, имаме оскъдни сведения за ранната история на ендемичните за Африка насекомоядни от разред Afrosoricida. Таралежоподобните тенреци, днес обитаващи Мадагаскар и ровещите златни къртици са първите известни бозайници от Миоцен за Източна Африка. Техните горни зъби се характеризират с V-образна форма (заламбдодонтизъм). Два от горните зъби на две все още некласифицирани животни от късен Палеоцен в Мароко са донякъде сходни със зъбите на тенреците и златните къртици и може би са техни предшественици. Но като се има предвид, че 33 млн. години делят тези две животни от тенреците и златните къртици, все още не са възможни каквито и да са заключения. Все пак трябва да имаме предвид, че тенреците и златните къртици са част от Afrotheria – голямото африканско разклонение на бозайниците, което включва също слоновете, африканския тръбозъб, слонската земеровка, даманите и морските крави. Слоновете се появяват първо през ранен Еоцен в Мароко, а групата им е много по-стара. Разделянето на различните групи от Afrotheria следователно е започнало поне през Палеоцен, от което следва, че е много вероятно Afrosoricida също да са древна група бозайници.
Освен видове, родствени на днешните насекомоядни, фауната през Палеоцен включва и подобни на тях плацентни бозайници, които изглежда принадлежат към самостоятелни групи. Тези групи биват причислявани към разред Proteutheria. Макар че е може би по-подходящо да бъдат разпределени в различни разреди – Leptictida, Pantolesta и Apatotheria.
Членовете на Leptictida първоначално са били считани за древни таралежи, което все още се представя в популярните реконструкции. Въпреки че някои учени продължават да причисляват тези животни към истинските насекомоядни, те всъщност са били напълно различни, особено с тенденцията задните им крайници да бъдат доста по-дълги от предните. Разредът се появява през късна Креда в западната част на Северна Америка с Gypsonictops – примитивни животни, обикновено характеризиращи се с пети предкътник, докато при повечето останали плацентни бозайници броят им е редуциран до четири. Разредът включва Leptictidae – предимно северноамериканско семейство, което възниква през ранен Палеоцен, представено от Prodiacodon и Palaeictops. Prodiacodon присъства до ранен Еоцен, докато Palaeictops се среща и до среден Еоцен и вероятно дава началото на Leptictis, като последният вид от този род е добре познат от намерените около 200 черепа в олигоценски слоеве. Дребните видове от Prodiacodon са били с размерите на мишка и тежащи около 30 грама, докато по-едрите от Prodiacodon и Palaeictops са достигали до над 200 грама (сравними с плъхове). Докато Prodiacodon са имали зъби с високи и заострени гънки, то при Palaeictops те стават все по-заоблени и ниски, свидетелствайки за преминаване към по-разнообразна диета.
 |
| Череп на Palaeictops. |
 |
Слонска земеровка. |
Докато Leptictida са обитавали горите на ранния Терциер, членовете на изчезналия разред Pantolesta изглежда са обитавали водите. Семейство Pantolestidae включва редица видове с все по-видроподобни адаптации, като започнем със среднопалеоценските Bessoecetor, минаваме през къснопалеоценските до ранноеоценски Palaeosinopa и стигнем до по-съвременните средноеоценски Pantolestes и Buxolestes. Bessoecetor и Palaeosinopa се появяват в Северна Америка, а Palaeosinopa се разпространява и в Европа в началото на Еоцен – периодът на фаунистичната размяна между северните континенти. Ранните пантолестиди от род Pagonomus обитават Европа от края на Палеоцен, което усложнява изводите за произхода на семейството. Пантолестидите достигат и Азия през Еоцен, като последният им представител преживява до началото на Олигоцен.
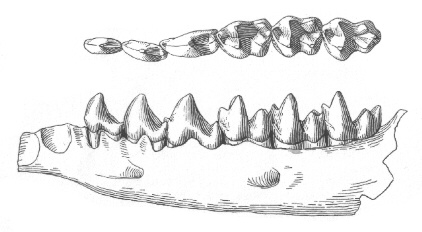 |
| Долна зъбна редица на Bessoecetor septentrionalis. |
Тъй като досега не е намерен скелет на пантолестид от Палеоцен, ние разчитаме на данните от еоценските представители. Отлично запазеният средноеоценски скелет в Месел, принадлежащ на пантолестида Buxolestes, е представен от два вида. По-големият от видовете има дължина на тялото и главата от 46 сантиметра и опашка дълга 35 сантиметра, което е много по-малко в сравнение с повечето съвременни видове видри. Предните и задните крайници са мощни, но без ясни специализации за плуване. Те завършват със здрави нокти, което може би е индикатор за способности за копаене на дупки. По-ясни белези за плувните навици на животните са открити в опашката: първият опашен прешлен има силно развити костни израстъци, за които вероятно са се закрепяли мощни мускули за задвижване на опашката, очевидно за генериране на изтласкващо движение във водна среда. Като последно потвърждение на полуводния начин на живот на Buxolestes при няколко индивида са открити фосилизирани останки от риба в стомашното си съдържимо. Намирани са останки и от други храни също така, но специално за популацията на Buxolestes в Месел няма данни за включване на миди и охлюви в диетата им, въпреки формата на зъбите им. В крайна сметка това най-вероятно са били животни, прекарващи много време във водата, ловейки риба, но много добре подвижни и на сушата също така.
Находките от Месел наскоро бяха допълнени от забележителен скелет на Palaeosinopa от ранния Еоцен на Уайоминг. Новият вид на този древен род пантолестиди е открит в скално образувание, което е известно с изобилните си фосилни находки на риби, а наскоро започнаха да се откриват и такива на бозайници. С дължина на тялото и главата 57 см това животно е по-близко по размери до днешните видри отколкото Buxolestes. Въпреки донякъде по-грацилната форма и по-дългата тънка опашка, скелетите на Palaeosinopa и Buxolestes са много сходни. И двата рода притежават основните адаптации за полуводен начин на живот. Като допълнение, стъпалата на Palaeosinopa са много големи и има вероятност да са били използвани при плуването. Фосилизираното стомашно съдържимо на Palaeosinopa показва, че подобно на Buxolestes и те са се хранили с риба.
 |
| Скелет на Palaeosinopa. |
Необичайна специализация е описана при Bisonalveus browni - малък пентакодонтид, тежащ около 25 грама. Доскоро фосилните находки от Bisonalveus се свеждаха до части от челюсти, но в Албърта, Канада бе открит почти цял череп, съдържащ първите известни кучешки зъби на Bisonalveus – те са дълги, с форма на кама, с характерна надлъжна бразда по предната страна на зъба. Подобни бразди се наблюдават в отровните зъби на съвременните змии, които им служат за провеждане на отровата. Очевидно дребничките Bisonalveus са притежавали подобен отровен апарат, който им е служил за обездвижване на плячката – предимно насекоми и други безгръбначни. Само за няколко вида съвременни бозайници, включително някои земеровки и соленодонът, насекомоядни обитаващи най-вече Карибските острови, е известно да притежават отровен апарат, като досега такъв не е бил откриван у изчезнали видове бозайници. Удивително е, че кучешки зъби на други, по-едри бозайници от късния Палеоцен на Албърта (все още неидентифицирани), също са притежават бразди, които са служили за извеждане на отрова. За разлика от отровните зъби на Bisonalveus тези зъби са от долната челюст и са със съвсем различно устройство, което показва, че отровният апарат еволюира независимо при тези две групи бозайници. Това означава, че отровните бозайници са били много по-разнообразна група през ранния Неозой, отколкото са днес.
 |
| Горен кучешки зъб и трети горен резец на Bisonalveus browni. |
Доколкото е известно, апатемидите са се срещали през ранен Терциер само в Северна Америка и Европа. Първите апатемиди се появяват през среден Палеоцен в района на Скалистите планини, представени от изключително примитивните Unuchinia и малко по-съвременните Jepsenella. В късен Палеоцен вторите биват заместени от Labidolemur. Докато видовете Unuchinia са достигали тегло от около 80 грама, то тези от Jepsenella и Labidolemur не са надвишавали размерите на мишка със своите 15 до 40 грама. При Labidolemur всъщност се развива устройство на зъбите, което характеризира семейството. Притежавали са по два горни резеца на всяка страна на челюстта (общо 4), първият от които е бил много уголемен и извит (като кука). Единственият долен резец (общо 2 за долната челюст) също е бил много уголемен, разположен напред и завършващ с извит нагоре и заострен край. Зъбите напомнят тези при гризачите. Всички останали резци и кучешки зъби липсват, също както и първите предкътници. Вторият предкътник е интересно видоизменен – неговата заострена коронка е издадена напред, покривайки част от основата на долните резци.
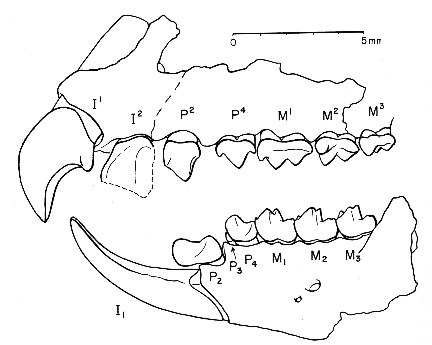 |
| Зъбна редица на Labidolemur kayi. |
 |
| Скелет на Apatemys chardini съхранен заедно с останки от рибата Knightia eocaenica. |
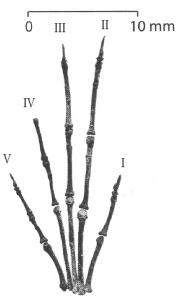 |
| Преден крайник на Labidolemur kayi. |
Насекомоядните бозайници са сред най-малко изучените представители на фауната в началото на Терциер. Но е ясно, че също както днешните насекомоядни, тези древни животни са притежавали забележителни специализации и са водели свой собствен, характерен начин на живот.
Изчезването на динозаврите освобождава пътя на плацентните бозайници и тяхното ограничено разнообразие в началото на Палеоцен е озадачаващо. Хищническиат натик от страна на динозаври често е спряган като причина за това, но тя не е така убедителна, когато говорим за малки бозайници. Насекомоядните оцеляват в свят пълен с хищници и днес, и е трудно да си представим, че динозаврите са представлявали по-голяма заплаха за тях, отколкото днешните хищници и грабливи птици за малките бозайници. Логично е да се заключи, че някои от екологичните ниши запълнени от насекомоядните бозайници, в края на креда са били заети от други групи животни. Малки хищни динозаври може би са се конкурирали с някои насекомоядни бозайници и вероятно са били лимитиращ фактор, като например насекомоядните птерозаври, особено дървесните. Но много по-важна конкуренция са представлявали торбестите, които са се характеризирали с висока численост и голямо разнообразие през късна Креда в Северна Америка, но почти изчезват от там през К/Т периода. Разбира се, в Южна Америка историята на бозайниците поема в друга посока, където торбестите вземат превес над насекомоядните в тези екологични ниши.
Няма коментари:
Публикуване на коментар