Никоя друга анатомична особеност не разграничава така красноречиво бозайниците и птиците, както перата и козината. Но така както двете групи се различават помежду си, се отличават и от всички останали гръбначни животни, благодарение на една обща физиологична особеност - способността да запазват постоянна телесна температура в покой, или ендотермия. Това често е ставало повод грешно да се допуска, че птиците и бозайниците имат общ произход.
Традиционно се смята, че развитието на космена покривка и пера, ускореният метаболизъм, както и способността за поддържане на относително постоянна температура на тялото, са тясно свързани. Приема се, че по-голямата способност за регулация на телесната температура при предшествениците на днешните бозайници, е резултат от едновременното развитие на повишени метаболитни процеси и изолационна космена покривка. По същия начин, тъй като ендотермната терморегулация се счита за неразривно свързана със способността за летене, се приема, че перата при предшествениците на днешните птици се развиват в отговор, както на нуждата от термоизолация, така и на летене. В действителност нито едно от тези предположения не съответства на наличните доказателства. Последни находки, фосилни и физиологически, показват, че повишената способност за терморегулация и при птици, и при бозайници, вероятно няма нищо общо с промените в метаболизма. По всичко изглежда, че перата и полетът при птиците са се появили много преди ендотермията.
Ендотермията е едно от най-големите еволюционни предимства, които се развиват при гръбначните животни, както и характеристика, отличаваща съвременните птици и бозайници от влечугите, земноводните и рибите. Ендотермията, която се развива независимо при бозайниците и птиците, представлява голямо предимство за организмите в екологично и физиологично отношение и може би е основна причина за днешния успех на тези две групи и широкото им разпространение в разнообразни водни и сухоземни екосистеми. Повишеният капацитет на белите дробове, потреблението на кислород, както и производството на вътрешна топлина (чрез аеробен метаболизъм), които са отличителни характеристики на ендотермията, позволяват на птици и бозайници да поддържат постоянна телесна температура в широк диапазон от температурни условия на околната среда. В резултат те могат да обитават местности с ниски или променливи температури, както и да водят нощен начин на живот, което е невъзможно за ектотермните животни. С малки изключения, ектотермните животни, като влечугите например, разчитат на анаеробни метаболитни процеси в случаите на бързо ускорително придвижване. Обикновено бързо настъпва умора, в резултат на натрупването на млечна киселина. Ендотермните животни от своя страна са в състояние да издържат сравнително високи нива на физическо натоварване за дълъг период от време.
Доскоро беше практически невъзможно да се докаже ендотермията при изчезнали животински видове. Ендотермията е процес, почти изцяло присъщ на меките тъкани, които оставят оскъдни или никакви вкаменелости. Физиологично ендотермията се постига чрез много високи нива на клетъчна консумация на кислород. В лабораторни условия тези показатели при бозайници в покой са обикновено 6 до 10 пъти по-високи от същите при влечуги със същото телесно тегло и температура на околната среда; показателите на птиците са дори още по-високи - около 15 пъти повече от тези на влечугите. В естествени условия метаболизмът на бозайниците и птиците често надвишава този при влечугите до 20 пъти. За да поддържат толкова високо равнище на консумация на кислород, при ендотермните животни настъпват големи анатомични и функционални промени за улесняване на транспорта и доставката на кислорода до тъканите. И при бозайници, и при птици се увеличава значително капацитета на белите дробове (при птиците и чрез т.нар. дивертикули), а двата кръга на кръвообръщението са напълно отделени. Повишава се обема на кръвта, заедно с по-високото потребление на кислород, както и завишената активност на тъканните ензими. Тези основни характеристики на ендотермната физиология е малко вероятно да бъдат съхранени във вкаменелости на птици, бозайници или други организми. Следователно предположенията за наличие на ендотермия при различни изчезнали гръбначни животни са хипотетични и се базират главно на съпоставянето между метаболитните процеси и някои косвени данни като съотношение хищник-жертва.
Съвременните проучвания за възможна ендотермия при изчезнали таксони наблягат на костната хистология и най-вече съпоставяне темповете на растеж на ектотермни и ендотермни животни. При съвременните гръбначни са установени два хистологични типа на изграждане на костите, различаващи се по фибрилната структура и степен на васкуларизация. Костите на съвременните земноводни и повечето влечуги спадат към така наречения ламеларно-зонален тип. Характерното за него е, че се наблюдават сравнително малко първични остеони в костната тъкан, които са свързани с образуването на костно вещество. В хистологично отношение ламеларно-зоналните кости имат стъпаловидна стуктура, с характерни кръгове на нарастване, а васкуларизацията е слаба. При фибро-ламеларните кости на птици, бозайници и динозаври в по-голямата част от костта се наблюдават обилно количество първични остеони, които произвеждат влакнеста костна тъкан. Фибро-ламеларните кости имат добра васкуларизация.
Ламеларно-зоналните кости се асоциират с ектотермията, а фибро-ламеларните - с ендотермията. Фибро-ламеларните кости са свързани с бързия растеж, което изисква интензивно отлагане на калциеви соли. Това интензивно нарастване е възможно само при организми с високи нива на метаболизъм, което пък се свързва с ендотермията. Т.е. основната връзка е между темповете на растеж и костната структура. Общоприето е, че темповете на растеж на съществуващите ендотермни животни в дивата природа са с порядък по-високи от тези на ектотермните. Като се има предвид, че повечето кости при динозаврите са фибро-ламеларни, може да се предположи, че темповете им на растеж са бързи, като при птиците и бозайниците. Което може да означава, че също като последните, динозаврите са били ендотермни животни.
Голяма част от тези предположения обаче са в противоречие с наличните данни от областта на палеонтологията и биологията. Например, известно е, че фибро-ламеларни кости са представени и в скелетите на днешните костенурки, крокодили и гущери. Освен това в дългите кости на много родове динозаври се наблюдават и двата хистологични типа костна тъкан - фибро-ламеларна и ламеларно-зонална.
Независимо от разликите в костната хистология, съществуват основания да се съмняваме в степента на различие в темповете на нарастване на ендотермните животни и влечугите, особено крокодилите, които са най-близките живи родственици на днешните птици и динозаврите. Според най-често цитираното сравнително изследване, регресията в темповете на нарастване (г/ден) при всички амниоти продължава до достигането на максималните размери при възрастните, като при влечугите този показател е по-висок със само 10% от този на ендотермните животни . Но критериите при изчисляването на регресията в нарастването на телесното тегло не са равностойни: ендотермните животни достигат максимално телесно тегло при полова зрялост и би било по-подходящо да се използва за сравнение този показател и при ектотермните в същия онтогенетичен етап, за по-голяма точност. Например, американският алигатор (Alligator mississippiensis) е посочен в изследването с максимално телесно тегло на възрастен индивид от 160 кг., което надвишава много теглото на индивидите в момента на достигане на полова зрялост - около 30 кг. На базата на тези данни нарастването на телесната маса на алигаторите е определено на 28 г/ден, а би било по-точно да е 42 г/ден. Когато бъдат взети предвид тези допълнителни данни, стойността на нарастване на телесното тегло при тези влечуги се оказва 4 пъти по-висока от тази на торбестите и много се доближава до стойностите на повечето плацентни бозайници. Особено важно е да се отбележи, че процентът на нарастване на телесното тегло при алигаторите е на практика неотличим от този на двукракият динозавър Troodon. Освен това Chinsamy и Dodson сравняват темповете на нарастване при три рода динозаври и не намират особени разлики (Chinsamy and Dodson, 1995).
От друга страна сравняването на съотношението на кислородните изотопи, О16:О18, във фосилизирани кости, показва макар и малки разлики в температурата на крайниците и вътрешната телесна температура при големите динозаври (в случая Nanotyrannus). Това показва, че тези големи динозаври са били ендотермни, тъй като съвременните ендотермни животни, за разлика от ектотермните, са способни да поддържат относително едни и същи стойности на периферната и вътрешната телесна температура. За съжаление известни са много противоречиви данни, които показват, че много птици и бозйници поддържат голяма разлика в периферната и вътрешната температура на тялото. Разбира се, възможно е съдържанието на кислородни изотопи във фосилизираните кости да е повлияно от условията на средата. Следователно съотношението на кислородните изотопи във фосилизирани динозавърски кости може да ни даде съвсем малко информация за метаболитната физиология на тези животни.
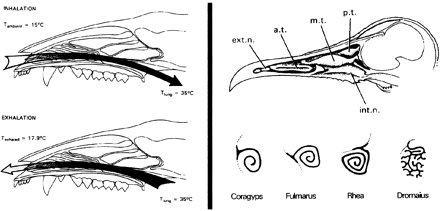
За разлика от по-горе представените факти, спиралната кост (в носната кухина) има силна функционална връзка с ендотермията. И при бозайниците, и при птиците ендотермията е тясно свързана с високи нива на консумация на кислород и съответно повишена вместимост на белите дробове. Спиралната кост, която се среща при 99% от всички бозайници и птици, улеснява обмена на топлина и влага между вдишвания въздух и лигавицата. По този начин тя значително ограничава загубата на влага и топлина, които иначе биха съпътствали увеличената вентилационна способност на белите дробове на ендотермните животни. Освен това отдавна се предполага, че появата при птици и бозайници на съдова мрежа между спиралната кост и мозъка се дължи на факта, че мозъкът използва тази система като охладител. Улесняването на охлаждането на мозъка би било особено важно при повишена телесна температура, или при интензивно физическо натоварване, когато се увеличава производството на вътрешна топлина.
Съществуващите бозайници, включително първичните яйцеснасящи (Prototheria) и живораждащите (Theria), вероятно произхождат от общ предшественик преди около 160 милиона години. Повечето бозайници поддържат сравнително постоянно ниво на метаболитни процеси и имат сходни анатомични особености с останалите ендотермни животни - космена покривка, ануклеарни еритроцити, белодробна структура, високо насищане на кръвта с кислород, диафрагма, потни жлези и т.н. Затова е логично да се предположи, че общият им мезозойски предшественик е притежавал подобна ендотермна физиология.
Изследването на физиологичния статус на по-древните бозайници, или протобозайници, се основава на фосилни свидетелства за техния метаболизъм. Бозайниците вероятно еволюират от цинодонтите или т.нар. бозайникоподобни влечуги в края на триас (преди около 200 милиона години) и отдавна се предполага, че физиология доближаваща се до ендотермната са притежавали още късните терапсиди.
Във фосилните находки на бозайници и бозайникоподобни влечуги наличието на назална спирална кост най-лесно се открива по костните ръбове, чрез които тя се прикрепя към стените на носната кухина. Разположението на прикрепящите ръбове на обонятелната спирална кост са разположени дорзално, далече от основния поток вдишан въздух, а тези на респираторната спирална кост в антеролатералната част, директно на пътя на вдишвания въздух. Костни ръбове за обонятелна спирална кост се срещат при всички бозайникоподобни влечуги, включително и примитивните пеликозаври. Докато елементарни антеролатерални ръбове за залавяне на респираторна спирална кост за първи път се появяват при някои големи, къснопалеозойски терапсиди, като Glanosuchus. Това означава, че началните етапи в развитието на бозайникоподобна кислородна консумация от организмите са започнали още в края на перм (преди около 250 милиона години) или 40 до 50 милиона години преди появата на бозайниците.
Наличието на елементарна спирална кост при пермските тероцефали е съществено за изучаването на еволюцията на ендотермията и космената покривка при предците на бозайниците. Моделите, свързващи произхода на космената покривка с началните етапи на развитие на ендотермия, са до голяма степен неточни, ако първите етапи на развитие на бозайников метаболизъм се наблюдават още при големите, къснопермски терапсиди. Много тероцефали изглежда са били активни хищници, достигащи размерите на куче, мечка или лъв (от 20 до 100 кг.) и са обитавали райони със субтропичен до тропичен климат. Няма доказателства, че са притежавали космена покривка. Но те са били достатъчно едри, така че независимо от типа на покривката, топлинната проводимост на телата им е съответствала на тази на едрите бозайници. Като се имат предвид климатичните пояси, които са обитавали, тероцефалите вероятно не са се нуждаели от специално изолационно покритие за поддържане на телесната температура. Следователно е много по-вероятно началните етапи на развитието на бозайников метаболизъм да са свързани с повишаването на кислородната консумация, отколкото с терморегулацията.
Цинодонтите от среден триас (напр. Thrinaxodon и Massetognathus), както и най-ранните бозайници (напр. Morganucodon) изглежда са притежавали ниво на развитие на респираторната спирална кост подобно на това на днешните бозайници. Следователно степента на белодробна вентилация, а оттам и метаболизма на ранните бозайници, по-специално на терапсидите от триас, е възможно да са достигали ниво на развитие като това на днешните бозайници. За част от тези ранномезозойски терапсиди се предполага, че са притежавали изолационна космена покривка, което се свързва със същинска ендотермна физиология. Доказателствата за козина при терапсидите се основават главно върху наличието на инфраорбитални ями и ръбове при някои триаски цинодонти. Предполага се, че те са свързани с наличието на косми със сензорна функция, а следователно и изолираща козина, която е покривала останалата част от тялото. Но ростралните отвори не свидетелстват задължително за наличието на сензорно окосмяване. Подобни образувания се намират и в черепите на ектотермни животни, по чиято люспеста кожа липсват сензорни косми или козина. При гущерът Tupinambis, например, ростралните отвори са почти еднакви по форма, брой и разположение с тези на цинодонтът Thrinaxodon. Във всеки случай е ясно, че сетивните косми по муцуната показват наличие на козина по тялото. Произходът на космената покривка не е добре изяснен, но ако първоначалната функция е била сензорна, е възможно сензорните косми да отразяват примитивизъм и да са се появили много преди цялостна изолационна космена покривка на тялото.
Изненадващо, най-новите данни сочат, че развитието на пълна космена покривка е невъзможно преди появата на най-ранните бозайници. Фосилни находки показват, че Хордеровата жлеза, която се свързва с наличието на козина при съвременните бозайници, се е срещала и при древните морганукодонти (напр. Morganucodon), но отсъства при терапсидите. За разлика от очните мукосерозни жлези на останалите четириноги, Хордеровата жлеза на бозайниците отделя предимно мастен секрет. Функциите на Хордеровата жлеза са проучени при гризачи, при които отделеният от нея мастен секрет се разнася по козината. Секретът на Хордеровата жлеза покрива козината и подобно на мастния секрет при птиците представлява бариера срещу студ, влага и определя отражателната способност на космената покривка.

По този начин наличните данни показват, въпреки все още да предстои потвърждение, че специализацията в назолакрималния комплекс при ранните бозайници може да се окаже свързана с трансформацията на Хордеровата жлеза в мастна и освобождаването на нейния секрет върху космената покривка. Това може би е свързано с появата на козината като изолационна покривка на тялото.
Възможно е изолационната космена покривка да става необходима за таксоните терапсидни бозайници, отличаващи се с малки размери (около 50 мм дължина на тялото) и нощен начин на живот. Може да се приеме, че космената изолационна покривка се развива при бозайниците във връзка с ендотермната регулация, но дълго след като бозайников тип метаболизъм вече е бил типичен за терапсидите.
Съвременните птици са ендотермни животни, като много от тях (особено пойните) поддържат най-висока телесна температура и високо ниво метаболитни процеси спрямо телесно тегло сред всички четириноги. Фосилни находки показват, че съвременните разреди птици са били определени още преди около 60 милиона години, ето защо ендотермната физиология на птиците може би е била вече добре представена през късна креда и началото на терциер.
Клас Птици е монофилетична група и не съществува родствен раннонеозойски таксон от който да може да се изведе логически появата на ендотермията (за разлика от отделните подгрупи при бозайниците - еднопроходни, торбести и плацентни). Най-ранната известна птица е Archaeopteryx lithographica, от късна юра (преди 145 милиона години). Тя притежава завършени крила и оперение на опашката, както и поразително сходство в някои отношения със скелети на месоядни динозаври. Въпреки това типът терморегулация е все още неизяснен. По-ранни косвени доказателства сочат, че Archaeopteryx вероятно е притежавал ектотермна физиология и много аспекти от анатомията и начинът на размножаване на съвременните птици свидетелстват, че способностите за летене предшестват ендотермията в еволюцията. Независимо от това, Archaeopteryx са понякога определяни условно като ендотермни , отчасти защото се предполага, че добре развитите им контурни пера са възпрепятствали достатъчно излъчването на телесна топлина в околната среда. Въпреки това, наличието на съвкупност от качества като летателна способност и оперение по тялото, и крилата в мезозойски предшественици на съвременните птици, не са задължително доказателство за наличието на ендотермия. Подобно на съвременните влечуги някои птици използват поведенчески методи за терморегулация и абсорбиране на топлина от околната среда през перушината. По време на студените нощи температурата на тялото на Geococcyx californianus (голяма земна кукувица) пада с около 4 градуса. След изгряване на слънцето земните кукувици излагат на припек по-слабо оперените части на тялото си и по този начин го затоплят, ектотермично. Освен това и други, напълно оперени птици, могат лесно да абсорбират и използват слънчева енергия. Ектотермни Archaeopteryx, които са обитавали местности с топъл и слънчев климат, е възможно да са имали сходни поведенчески механизми за терморегулация. Напълно оперените Archaeopteryx, независимо дали ектотермни или ендотермни, вероятно лесно са поддържали постоянно ниво на телесна температура. Следователно появата на добре развита перушина толкова рано в еволюцията на птиците е по-правилно да се свърже с усъвършенстването и развитието на летателните им способности, отколкото с терморегулацията.
Анатомичните сходства между някои мезозойски архозаври и Archaeopteryx са факт и много учени ги приемат като доказателство за близкородствена връзка между дромеозаврите (като Velociraptor и Deinonychus) и примитивните птици. Следоватлено метаболитният статус на тези динозаври може да предостави ценна информация относно екто- или ендотермната физиология на ранните птици.
Наличието или отсъствието на ендотермия при динозаврите е тема, представляваща обект на понякога дори ожесточени дискусии през последните повече от 20 години. Ясно е, че големите динозаври са имали пасивна терморегулация. Но както беше отбелязано по-горе, някои данни (като съотношение хищник-плячка, поза на тялото, костна хистология) свидетелстват недвусмислено в подкрепа на ендотермия.
Определянето на наличие или липса на дихателна спирална кост при тероподите може да осигури най-прякото доказателство за метаболитния им статус. Но няколко фактора усложняват изследването на еволюцията на спиралната кост при птиците и техните предшественици. Въпреки че спиралната кост се калцифицира и вкостява, при много от съществуващите таксони тази структура често остава хрущялна, с липса на характерните костни места за захващане в носната кухина, което значително намалява шансовете за директно доказване на наличието или липсата и в изчезнали таксони. Въпреки това е установено, че наличието на дихателна спирална кост при днешните ендотермни животни е свързано с пропорционалното разширяване на напречното сечение на носната кухина. Това е във връзка както с увеличената вентилация на белите дробове, така и с доставянето на по-голям обем въздух в спиралната кост. Показателно е, че относителната ширина на носната кухина при все по-късните и бозайникоподобни терапсиди нараства и се доближава до, а при Thrinaxodon дори достига, тази на бозайниците и птиците.
Най-нови методи с използването на компютърна томография улесниха много неинвазивното детайлно изследване на носната кухина при фосилизирани екземпляри. При Tyrannosaurus сканирането ясно показва, че е малко вероятно тези животни да са притежавали респираторна спирална кост: такива кости отсъстват от изкопаемите останки и, което е по-важно, напречното сечение на носната кухина е идентично с това на днешните ектотермни животни. Освен това компютърното сканиране на носните кухини и на други тероподни динозаври, Ornithomimus и Hypacrosaurus, също показва, че са били твърде тесни и с недостатъчен обем за разполагането на спирална кост в тях. Тези характеристики са изключително сходни с данните при съвременните влечуги (напр. крокодилите) и е силно доказателство за ниското ниво на белодробна вентилация и ектотермия при тези динозаври.
Тесните назални пътища при тероподите са свързани с наличието на допълнителни обемни пневматични синуси и техните отличителни външни отвори. За разлика от тях, при днешните птици липсват допълнителни анторбитални синуси, защото техните широки назални пътища са почти запълнени от респираторна спирална кост и няма достатъчно място за допълнителни синуси. Забележително е, че Archaeopteryx притежава анторбитални отвори и широки допълнителни синуси, подобно на тероподите. Следователно, подобно на своите предшественици, тероподните динозаври, е малко вероятно Archaeopteryx да е притежавал респираторна спирална кост или ендотермен метаболитен статус. За ввсеки случай мезозойският климат е бил толкова мек и благоприятен, че дори ранноюрските птици са били в състояние лесно да поддържат относително постоянно ниво на терморегулация.

Следователно е най-логично да се приеме, че перата са се развили във връзка с летенето, а не с терморегулацията. Както посочва Feduccia (1996), перата са най-сложните рогови образувания на епидермиса, при всички животни те са разположени аеродинамично, а контурните и маховите пера имат тенденцията да дегенерират при вторично нелетящите видове. Много е вероятно ендотермията при птиците да еволюира дълго след развиване на летателните им способности, като начин за подобряване на издържливостта при дълъг полет, а не за регулиране на температурата.
Неотдавна описани фосилни находки на раннокредни влечуги в Китай (провинция Ляонин) се предполага, че представляват пернати тероподни динозаври. Тези твърдения следва да се разглеждат с повишено внимание. Компсогнатусът Sinosauropteryx притежава по средната линия на гърба подобни на косми влакна, които се простират по протежение на врата, гърба и опашката. Първоначалните заключения, че тези влакна най-вероятно представляват протопера, всяко от които се състои от централна ос и разклонения, все още не са потвърдени. Подробното изследване на тези влакна показва, че е много вероятно да представляват мацерирали колагенови фибри. Тези данни съответстват с наличието на такива влакна и при други тероподи в Китай.

Други два рода, Protarchaeopteryx и Caudipteryx, притежават добре обособени пера, но статусът им на тероподи е съмнителен. Има твърде малко останки от Protarchaeopteryx, за да се направи окончателен таксономичен анализ. Въпреки това дългите предни крайници не са в противоречие с родството с птиците. Caudipteryx притежава къси, оперени предни крайници и е много вероятно да представлява вторично нелетяща птица. Основанията за класифицирането му като теропод се базират главно на три недвусмислени анатомични особености, които го определят като протоптица. Те включват: (1) срастване в областта на костите на долната челюст - квадратоюгална и квадратна - или липса на последната; (2) контакт на квадратоюгалната кост със слепоочната; (3) израстък на седалищната кост, срастващ се с пубиса. Въпреки това, при Caudipteryx тези особености или не съществуват, или са твърде неясни. По-рано публикувани илюстрации сочат, че няма контакт между квадратоюгалната и квадратна кост, а квадратоюгалната е твърде къса за свързване със сквамозата. В крайна сметка израстъкът на седалищната кост може да съществува или не при Caudipteryx, но със сигурност подобна структура се наблюдава при някои птици.

Развитието на ендотермен метаболизъм и козина и пера при ранните бозайници и птици съответно, имат коренно различна хронология. Наличието на добре развита респираторна спирална кост при среднотриаски цинодонти показва, че достигането на високи, бозайникоподобни нива на метаболитни процеси вероятно предшества произхода на бозайниците с поне 10-15 милиона години. Въпреки това, най-ранните изкопаеми данни за развитието на Хордерови жлези показват, че развитие на космена покривка не се среща преди появата на бозайниците. Много е вероятно малките размери и нощният начин на живот на първите бозайници също да са изиграли важна роля за развитието на изолационна космена покривка за подпомагане на терморегулацията.
Традиционно се приема, че липсата на респираторна спирална кост при тероподните динозаври и Archaeopteryx е силно доказателство в подкрепа на схващането, че ранните птици са били ектотермни. Така че появата на пера и способности за летене вероятно предхождат развитието на ендотермия при птиците. Най-логично е да предположим, че перата се появяват в отговор на нуждата от ареодинамика, а не терморегулация.
Няма коментари:
Публикуване на коментар